Генетика бактерий и вирусов. Генетический материал бактерий. Нуклеоидная мутация –
происходящая в нуклеоиде
Генетика микроорганизмов как наука
Замечание 1
Примерно до конца $30$-х годов $XX$ века считалось, что микроорганизмы не имеют ядерного аппарата. Поэтому вопросы наследственности и изменчивости микроорганизмов тщательно не изучались.
Лишь с изобретением электронного микроскопа появилась возможность рассмотреть субмикроскопическую структуру клетки вообще и микроорганизмов в частности.
С начала $40$-х годов ученые-генетики обращают свое внимание на микроорганизмы. Бактерии, микроскопические грибы и вирусы становятся объектами генетических исследований. Формируется новая отрасль микробиологии – генетика микроорганизмов.
Генетика микроорганизмов – это раздел общей генетики, в котором предметом изучения служат микроорганизмы (бактерии, вирусы, микроскопические грибы) и особенности их наследственности и изменчивости.
Характерной особенностью микроорганизмов является гаплоидный набор хромосом или кольцевая молекула ДНК. Это дает возможность мутациям проявиться уже в первом поколении потомков.
Начало микробиологических генетических исследований
Благодаря изучению субмикроскопической структуры клеток микроорганизмов удалось найти ответы на многие вопросы генетики. Американсие генетики О.Т. Эйвери, К. Мак-Леод и М. Маккарти, проводя опыты на пневмококках, получили первые доказательства того, что материальным носителем наследственности является молекула ДНК. Исследования хлебной плесени позволило сформулировать положение, что один ген программирует синтез одной полипептидной цепи (одногобелка).
Но особенно интенсивно стали исследовать микроорганизмы с точки зрения генетики после того, как американскими микробиологами С. Лурия и М. Дельброком на примере кишечной палочки было доказано универсальность закономерностей мутационного процесса. Они доказали, что и бактерии подчиняются мутационным закономерностям.
В науке появился новый принцип изучения изменчивости у бактерий – клональный анализ. Он заключается в тщательном исследовании потомства одной клетки. Эта клетка становится родоначальником клона.
Изучение бактерий
В результате кропотливых исследований американским генетикам Дж. И Э. Ледербергам удалось доказать, что у бактерий мутации возникают независимо от условий их культивирования. Они разработали метод отпечатков, который позволил очень упростить приемы отбора микроорганизмов с желаемыми свойствами для дальнейших исследований. Они доказали, что больших популяциях клеток бактерий мутации происходят неупорядочено – спонтанно.
В $1946$ году было доказано, что бактериям тоже присущ половой процесс, были открыты явления конъюгации хромосом и рекомбинации генов, переноса генетической информации от одной бактериальной клетки к другой при посредстве бактериофага.
Существует мнение, что в кольцевой молекуле нуклеиновой кислоты клеток прокариот «прочтение информации» зависит от места начала «считывания». В зависимости от того, с какого нуклеотида начался этот процесс, находится и синтез того или иного белка.
Изучение фагов
Изучая особенности взаимоотношений «бактерия – бактериофаг», американские генетики открыли явление трансдукции (переноса генов между бактериальными клетками с помощью фагов) и обнаружили рекомбинацию у фагов. Это дало возможность изучать вопросы наследственности на уровне молекул (молекулярный уровень организации материи).
Немецкие микробиологи исследовали молекулу РНК. Для каждой из групп микроорганизмов была разработана методика исследований.
Генетика грибов и водорослей
Низшие грибы и водоросли имеют половой процесс несколько отличный от полового процесса других организмов. Благодаря их изучению появился новый метод – тетрадный анализ. Исследуя эти организмы, ученые разрабатывали методику объединения ядер генетически различных штаммов микроорганизмов. Все эти методы могут в дальнейшем послужить для выведения организмов с заданными качествами, для разработки новых поколений антибиотиков и биологически активных веществ, а также для борьбы со многими видами заболеваний растений, животных и, конечно же, человека.
Замечание 2
Но вопросы генной инженерии требуют осторожного подхода к изучению и применению полученной информации на практике. Ведь не ясно, к каким последствиям может привести появление генетически модифицированных организмов в природе и в человеческом организме.
Данная книга предназначена студентам медицинских образовательных учреждений. Это краткое пособие поможет при подготовке и сдаче экзамена по микробиологии. Материал изложен в очень удобной и запоминающейся форме и поможет студентам за сжатый срок детально освоить основные концепции и понятия курса, а также конкретизировать и систематизировать знания.
* * *
Приведённый ознакомительный фрагмент книги Микробиология: конспект лекций (К. В. Ткаченко) предоставлен нашим книжным партнёром - компанией ЛитРес .
ЛЕКЦИЯ № 4. Генетика микроорганизмов. Бактериофаги
1. Организация наследственного материала бактерий
Наследственный аппарат бактерий представлен одной хромосомой, которая представляет собой молекулу ДНК, она спирализована и свернута в кольцо. Это кольцо в одной точке прикреплено к цитоплазматической мембране. На бактериальной хромосоме располагаются отдельные гены.
Функциональными единицами генома бактерий, кроме хромосомных генов, являются:
1) IS-последовательности;
2) транспозоны;
3) плазмиды.
IS-последовательности – это короткие фрагменты ДНК. Они не несут структурных (кодирующих белок) генов, а содержат только гены, ответственные за транспозицию (способность перемещаться по хромосоме и встраиваться в различные ее участки).
Транспозоны – это более крупные молекулы ДНК. Помимо генов, ответственных за транспозицию, они содержат и структурный ген. Транспозоны способны перемещаться по хромосоме. Их положение сказывается на экспрессии генов. Транспозоны могут существовать и вне хромосомы (автономно), но неспособны к автономной репликации.
Плазмиды – дополнительный внехромосомный генетический материал. Представляет собой кольцевую, двунитевую молекулу ДНК, гены которой кодируют дополнительные свойства, придавая селективные преимущества клеткам. Плазмиды способны к автономной репликации, т. е. независимо от хромосомы или под слабым ее контролем. За счет автономной репликации плазмиды могут давать явление амплификации: одна и та же плазмида может находиться в нескольких копиях, тем самым усиливая проявление данного признака.
В зависимости от свойств признаков, которые кодируют плазмиды, различают:
1) R-плазмиды. Обеспечивают лекарственную устойчивость; могут содержать гены, ответственные за синтез ферментов, разрушающих лекарственные вещества, могут менять проницаемость мембран;
2) F-плазмиды. Кодируют пол у бактерий. Мужские клетки (F+) содержат F-плазмиду, женские (F-) – не содержат. Мужские клетки выступают в роли донора генетического материала при конъюгации, а женские – реципиента. Они отличаются поверхностным электрическим зарядом и поэтому притягиваются. От донора переходит сама F-плазмида, если она находится в автономном состоянии в клетке.
F-плазмиды способны интегрировать в хромосому клетки и выходить из интегрированного состояния в автономное. При этом захватываются хромосомные гены, которые клетка может отдавать при конъюгации;
3) Col-плазмиды. Кодируют синтез бактериоцинов. Это бактерицидные вещества, действующие на близкородственные бактерии;
4) Tox-плазмиды. Кодируют выработку экзотоксинов;
5) плазмиды биодеградации. Кодируют ферменты, с помощью которых бактерии могут утилизировать ксенобиотики.
Потеря клеткой плазмиды не приводит к ее гибели. В одной и той же клетке могут находиться разные плазмиды.
2. Изменчивость у бактерий
Различают два вида изменчивости – фенотипическую и генотипическую.
Фенотипическая изменчивость – модификации – не затрагивает генотип. Модификации затрагивают большинство особей в популяции. Они не передаются по наследству и с течением времени затухают, т. е. возвращаются к исходному фенотипу.
Генотипическая изменчивость затрагивает генотип. В основе ее лежат мутации и рекомбинации.
Мутации – изменение генотипа, сохраняющееся в ряду поколений и сопровождающееся изменением фенотипа. Особенностями мутаций у бактерий является относительная легкость их выявления.
По локализации различают мутации:
1) генные (точечные);
2) хромосомные;
3) плазмидные.
По происхождению мутации могут быть:
1) спонтанными (мутаген неизвестен);
2) индуцированными (мутаген неизвестен).
Рекомбинации – это обмен генетическим материалом между двумя особями с появлением рекомбинантных особей с измененным генотипом.
У бактерий существует несколько механизмов рекомбинации:
1) конъюгация;
2) слияние протопластов;
3) трансформация;
4) трансдукция.
Конъюгация – обмен генетической информацией при непосредственном контакте донора и реципиента. Наиболее высокая частота передачи у плазмид, при этом плазмиды могут иметь разных хозяев. После образования между донором и реципиентом конъюгационного мостика одна нить ДНК-донора поступает по нему в клетку-реципиент. Чем дольше этот контакт, тем большая часть донорской ДНК может быть передана реципиенту.
Слияние протопластов – механизм обмена генетической информацией при непосредственном контакте участков цитоплазматической мембраны у бактерий, лишенных клеточной стенки.
Трансформация – передача генетической информации в виде изолированных фрагментов ДНК при нахождении реципиентной клетки в среде, содержащей ДНК-донора. Для трансдукции необходимо особое физиологическое состояние клетки-реципиента – компетентность. Это состояние присуще активно делящимся клеткам, в которых идут процессы репликации собственных нуклеиновых кислот. В таких клетках действует фактор компетенции – это белок, который вызывает повышение проницаемости клеточной стенки и цитоплазматической мембраны, поэтому фрагмент ДНК может проникать в такую клетку.
Трансдукция – это передача генетической информации между бактериальными клетками с помощью умеренных трансдуцирующих фагов. Трансдуцирующие фаги могут переносить один ген или более.
Трансдукция бывает:
1) специфической (переносится всегда один и тот же ген, трансдуцирующий фаг всегда располагается в одном и том же месте);
2) неспецифической (передаются разные гены, локализация трансдуцирующего фага непостоянна).
3. Бактериофаги
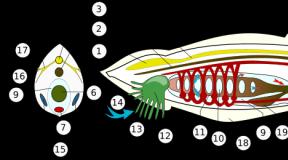
Вирионы фагов состоят из головки, содержащей нуклеиновую кислоту вируса, и отростка.
Нуклеокапсид головки фага имеет кубический тип симметрии, а отросток – спиральный тип, т. е. бактериофаги имеют смешанный тип симметрии.
Фаги могут существовать в двух формах:
1) внутриклеточной (это профаг, чистая ДНК);
2) внеклеточной (это вирион).
Фаги, как и другие вирусы, обладают антигенными свойствами и содержат группоспецифические и типоспецифические антигены.
Различают два типа взаимодействия фага с клеткой:
1) литический (продуктивная вирусная инфекция). Это тип взаимодействия, при котором происходит репродукция вируса в бактериальной клетке. Она при этом погибает. Вначале происходит адсорбция фагов на клеточной стенке. Затем следует фаза проникновения. В месте адсорбции фага действует лизоцим, и за счет сократительных белков хвостовой части в клетку впрыскивается нуклеиновая кислота фага. Далее следует средний период, в течение которого подавляется синтез клеточных компонентов и осуществляется дисконъюнктивный способ репродукции фага. При этом в области нуклеоида синтезируется нуклеиновая кислота фага, а затем на рибосомах осуществляется синтез белка. Фаги, обладающие литическим типом взаимодействия, называют вирулентными.
В заключительный период в результате самосборки белки укладываются вокруг нуклеиновой кислоты и образуются новые частицы фагов. Они выходят из клетки, разрывая ее клеточную стенку, т. е. происходит лизис бактерии;
2) лизогенный. Это умеренные фаги. При проникновении нуклеиновой кислоты в клетку идет интеграция ее в геном клетки, наблюдается длительное сожительство фага с клеткой без ее гибели. При изменении внешних условий могут происходить выход фага из интегрированной формы и развитие продуктивной вирусной инфекции.
По признаку специфичности выделяют:
1) поливалентные фаги (лизируют культуры одного семейства или рода бактерий);
2) моновалентные (лизируют культуры только одного вида бактерий);
3) типовые (способны вызывать лизис только определенных типов (вариантов) бактериальной культуры внутри вида бактерий).
Фаги могут применяться в качестве диагностических препаратов для установления рода и вида бактерий, выделенных в ходе бактериологического исследования. Однако чаще их применяют для лечения и профилактики некоторых инфекционных заболеваний.
Оглавление темы "Оценка роста бактерий. Спорообразование бактериями. Генетика бактерий.":1. Двухфазный рост бактерий. Диауксия. Рост без деления. Оценка роста бактерий. Количественная оценка роста бактерий.
2. Факторы влияющие на рост бактерий. Культуральные среды для роста бактерий. Простые и сложные культуральные среды. Твердые и жидкие культуральные среды.
3. Температура роста бактерий. Мезофильные бактерии. Термофильные бактерии. Психрофильные бактерии. Аэрация бактерий.
4. Величина рН необходимая для роста бактерий. Пигменты бактерий. Виды пигментов. Функции пигментов бактерий.
5. Спорообразование бактериями. Споры бактерий. Спорангии. Эндоспоры. Экзоспоры.
6. Морфология споры бактерий. Строение споры бактерий. Структура споры бактерий.
7. Споруляция бактерий. Стадии споруляции у бактерий. Расположение спор у бактерий.
8. Прорастание спор бактерий. Активация споры. Покоящиеся (некультивируемые) формы бактерий.
10. Внехромосомные факторы наследственности бактерий. Плазмиды бактерии. Виды плазмид. Функции плазмид бактерий.
В наши дни приоритетным направлением естествознания можно считать молекулярную биологию. Она тесно связана с микробиологией и в известном смысле является её детищем, так как в качестве основных моделей использует бактерии и вирусы, а одно из основных направлений молекулярной биологии - молекулярная генетика - долгое время являлась не чем иным, как генетикой бактерий и бактериофагов.
Изучение генетики бактерий имеет также и несомненный прикладной интерес, например в плане установления механизмов передачи патогенных свойств и устойчивости к лекарственным препаратам.
Бактерии - удобная модель для генетических исследований . Их отличает: относительная простота строения генома, позволяющая выявлять мутанты с частотой 10 -9 и ниже; гаплоидность, исключающая явление доминантности; половая дифференциация в виде донорских и реципиентных клеток; наличие обособленных, и интегрированных фрагментов ДНК (плазмид, транспозонов и т.д.); лёгкость культивирования и возможность получения популяций, содержащих миллиарды микробных тел.
Как и у других организмов, совокупность генов бактериальной клетки - геном - определяет её свойства и признаки (генотип ). Фенотип бактериальной клетки - результат взаимодействий между бактерией и окружающей средой - также контролирует геном (так как сами признаки закодированы в бактериальных генах).
Генетический материал бактерий
Ядерные структуры бактерий имеют характерное строение, отличающее их от ядер эукарио-тических клеток; их образуют так называемые хроматиновые тельца, или нуклеоиды, лишённые оболочки и включающие в себя почти всю ДНК бактерии.
Ядерные структуры можно наблюдать в фазово-контрастный микроскоп, где они выглядят как менее плотные участки цитоплазмы. Для их выявления в фиксированных мазках предложена реакция Фёльгена-Россенбёка.
В растущих бактериальных клетках нуклеоиды активно делятся, их количество иногда достигает 2-4.

Прокариотический геном
У бактерий обычно имеется одна замкнутая кольцевидная хромосома , содержащая до 4000 отдельных генов, необходимых для поддержания жизнедеятельности и размножения бактерий, то есть бактериальная клетка гаплоидна, а удвоение хромосомы обычно сопровождается её делением.
Некоторые виды (например, Brucella melitensis) стабильно содержат две кольцевые хромосомы , другие (Leptospira interrogans) - одну кольцевую хромосому и одну большую плазмиду, третьи - одну линейную хромосому (Streptomyces ambofaciens), то есть обладают сложными геномами.
Бактериальная хромосома содержит до 5*10 6 пар оснований. Для сравнения: геном человека составляет 2,9*10 9 пар оснований. Длина бактериальной хромосомы в развёрнутом состоянии составляет около 1 мм (Escherichia coli).
Некоторые бактерии содержат внехромосомные молекулы ДНК (плазмиды ) и мобильные элементы (либо плазмидные, либо хромосомные).
ЛФ, ФИУ, ПФ. Занятие № 6
А. Основные положения
Организация генетического материала у бактерий.
Наследственная информация бактерий хранится в ДНК, которая в прокариотической клетке является циркулярно замкнутой, двух цепочечной, суперспирализованной и представлена двумя типами молекул: большая – нуклеоид, где закодированы жизненно важные признаки, и малые – внехромосомные факторы наследственности (плазмиды, транспозоны, IS-последовательности, и умеренные бактериофаги), в которых закодированы дополнительные признаки.
Внехромосомные факторы наследственности и их встраивание их в нуклеоид.
Плазмиды могут, подобно нуклеоиду, самореплицироваться и поэтому относятся к автономным факторам наследственности (в отличие от остальных – неавтономных, которые способны реплицироваться лишь в составе нуклеоида или плазмиды), кроме того, плазмиды, как и умеренные фаги, могут встраиваться в нуклеоид только в гомологичных участках, в отличие от транспозонов и IS-последовательностей, способных встраиваться в нуклеоид в любых его участках.
Плазмиды.
Плазмиды выполняют в бактериальной клетке две возможные функции – регуляторную (содержа в себя дупликаты некоторых нуклеоидных генов) и кодирующую (неся гены, которых в нуклеоиде нет), могут существовать в двух состояниях (автономном, т.е. вне нуклеоида, и интегрированном в нуклеоид), а в зависимости от содержания в них tra-оперона – быть конъюгативными (если этот оперон содержится в данной плазмиде) и неконъгативными.
Функции tra-оперона.
Tra-оперон обуславливает возможность процесса конъюгации (детерминирует образование конъюгативных пилей и процесс одностороннего переноса через них генетического материала: плазмиды или участка нуклеоида).
F-плазмиды.
F-плазмиды представляют собой собственно tra-оперон, без каких-либо дополнительных генов, они детерминирует перенос при конъюгации себя, а также другую молекулу ДНК, в которую интегрирована F-плазмида (если это молекула ДНК – неконъюгационная плазмида, то в результате интеграции F-плазмиды она становится конъюгационной и передаётся через конъюгационную пилю, если это молекула ДНК – нуклеоид, то при конъюгации передаётся его часть, но не сама F-плазмида).
Формирование различных состояний плазмиды F.
F-плазмида из автономного состояния может переходить в интегрированное в нуклеоид состояние (в этом случае она называется Hfr-фактором), а также возвращаться в автономное состояние (или полностью сохраняя свой состав или «обмениваясь» с нуклеоидом своим концевым участком – в последнем случая такая рекомбинантная плазмида называется F’-плазмидой).
R-плазмиды.
R-плазмиды – это плазмиды, детерминирующие множественную лекарственную устойчивость (или резистентность, откуда и название) бактериальной клетки к антибиотикам; они состоят из генов, детерминирующих такую устойчивость (r-оперон) и F-плазмиды (которая в этом случае носит название RTF-фактора.
Состав r-оперона.
Этот оперон содержит гены, детерминирующие устойчивость к антибиотикам, а также в его состав может входить транспозон или его часть – IS-последовательность.
Механизмы устойчивости бактерий к антибиотикам, обусловленные наличием R-плазмид.
R-плазмида может детерминировать: способность бактериальной клетки инактивировать антибиотик, способность бактериальной клетки модифицировать антибиотик с потерей последним своей антибактериальной активности, способность бактериальной клетки снижать проницаемость клеточной стенки для данного антибиотика.
Бактериоциногенные плазмиды (на примере col-плазмиды E.coli).
Бактериоциногенные плазмиды детерминируют синтез антибиотикоподобных веществ – бактериоцинов; у кишечной палочки бактериоциногенные плазмиды называются колициногенными (как и у других бактерий – по названию вида) или Col-плазмидами, а бактериоцины – колицинами (по такому же принципу).
Характеристика колицинов.
Колицины представляют собой белки, которые убивают бактериальную клетку, но не лизируют её.
Транспозоны.
Транспозоны – нуклеотидные последовательности, включающие в себя IS-последовательности (которые обуславливают способность транспозонов менять место локализации в молекуле ДНК, а также «перескакивать» из одной молекулы ДНК в другую – так называемые «прыгающие гены»), гены, детерминирующие какой-либо признак, а также особые концевые структуры, благодаря которым транспозоны могут находиться в автономном состоянии (поскольку, благодаря этим «липким концам» замыкаются в кольцо).
IS-последовательности.
IS-последовательности включают в себя только гены транспозиции, в отличие от транспозонов не могут находиться в автономно состоянии.
Механизмы фаговой конверсии.
Когда в результате лизогенизации (т.е. встраивание в геном умеренного бактериофага) бактерия приобретает дополнительный признак, это может быть обусловлено тремя возможными механизмами: дерепрессией гена профага, внесением соответствующего гена дефектным фагом, запуском «молчащего» из-за повреждения собственного промотора гена промотором профага.
Модификации у бактерий.
Модификациями называются фенотипическая изменчивость у бактерий.
Мутации у бактерий.
При мутационной изменчивости у бактерий происходят изменения в первичной структуре ДНК, которые выражаются в наследственно закреплённой утрате или изменении какого-либо признака (или признаков); при спонтанных мутациях факторы, их вызвавший (мутаген) не известен – наиболее часто спонтанные мутации происходят в результате ошибки ДНК-полимеразы при репликации ДНК, среди спонтанных мутаций выделяют инсертационные мутации, которые происходят вследствие встраивания в нуклеоид внехромосомных факторов наследственности; индуцированные мутации у бактерий вызываются в эксперименте действием конкретного мутагена.
SR-диссоциации.
SR-диссоциацией называется такое явление, когда в чистой культуре, образующей S-формы колоний, появляются R-формы; по своему механизму SR-диссоциация – это инсертационная мутация, приводящая к утрате генов, контролирующих синтез полисахаридных звеньев ЛПС наружной мембраны клеточной стенки.
Мутагены.
Под мутагенами понимают химические вещества или физические факторы, вызывающие предмутационные изменения в ДНК, которые в результате ошибок репарирующих ферментов или в процессе репарации переходят в мутацию.
Репарации у бактерий.
Этим термином обозначают процесс восстановления повреждённой ДНК ферментами репарационных систем бактериальной клетки.
Рекомбинационная изменчивость у бактерий.
Под рекомбинационной изменчивостью понимают изменчивость, происходящую в результате включения в ДНК реципиентной клетки участка ДНК донорской клетки; у бактерий насчитывают пять видов генетических рекомбинаций (т.е. пять видов рекомбинационной изменчивости): трансформация (непосредственная передача генетического материала от донорской к реципиентной клетке), трансдукция (передача генетического материала от донорской к реципиентной клетке с помощью дефектных бактериофагов), конъюгация (передача генетического материала от донорской к реципиентной клетке с помощью конъюгационных пилей), лизогенизация (при ней в геном реципиентной клетки внедряется экзогенный генетический материал – геном умеренного фага), фаговая конверсия (отличается от лизогенизации лишь тем, что фенотип донорской клетки изменяется).
Генная инженерия в медицинской микробиологии.
В медицинской микробиологии все шире используются методы генной инженерии, с помощью которых «заставляют» микроорганизмы продуцировать нужные медицинской практике препараты (вакцины, гормоны, интерфероны, цитокины и др.), путем внесения в их геном соответствующего гена, т.е. получения рекомбинантного штамма с нужными свойствами путем «направленной» рекомбинационной изменчивости.
Генетические методы, применяемые в микробиологической диагностике.
В современной медицине все большее распространение получают генетические методы микробиологической диагностики: определение процентного содержания гуанина и цитозина в бактериальном геноме, метод молекулярной гибридизации и особенно – полимеразная цепная реакция (ПЦР).
Метод молекулярной гибридизации.
Этот метод используется для выявления степени сходства различных ДНК (при идентификации микроорганизмов проводят сравнение ДНК выделенного штамма с ДНК эталонного штамма).
Полимеразная цепная реакция.
ПЦР можно проводить для достижения трех целей: для обнаружения в патологическом материале конкретного вида микроорганизма без выделения чистой культуры, для идентификации выделенных чистых культур микроорганизмов, для генотипирования микроорганизмов, т.е. определения генетических вариантов одного вида; принцип осуществления ПЦР заключается в увеличении (амплификации) количества искомого гена при положительной или отсутствии такого увеличения при отрицательной реакции (т.е. проводится экстракция ДНК и, если в ней содержится искомый ген, ее количество в процессе реакции резко увеличивается, что выявляется с помощью электрофореза).
Б. Лекционный курс









































В. Теоретический материал
| 11. Организация генетического материала у бактерий | |
| 11.2. Плазмиды | |
| 11.3. F-плазмиды | |
| 11.4. R-плазмиды | |
| 11.6. Транспозоны | |
| 11.7. IS-последовательности | |
| 12. Модификации, мутации и репарации у бактерий | |
| 12.1. Модификации у бактерий | |
| 12.2. Мутации у бактерий | |
| 12.3. SR-диссоциация | |
| 12.4. Мутагены | |
| 12.5. Репарации | |
| 13. Генетические рекомбинации у бактерий, генная инженерия в медицинской микробиологии | |
| 13.1. Виды генетических рекомбинаций у бактерий | |
| 13.2. Генная инженерия в медицинской микробиологии | |
| 14. Применение генетических методов в микробиологической диагностике | |
| 14.1. Метод молекулярной гибридизации | |
| 14.2. Полимеразная цепная реакция |
ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА У БАКТЕРИЙ
11.1. Общая схема организации генетического материала бактериальной клетки
Организация генетического материала у бактерий основана на общем для всех форм на Земле принципе. Поэтому данный материал излагается с учетом знания студентами курса общей генетики: освещаются лишь те генетические особенности, которые характерны для прокариотической клетки. Наследственная информация бактерий, как и у всех форм клеточной жизни (в отличие от вирусов), хранится в ДНК (Рис. 11.1-1), которая в прокариотической клетке представлена двумя типами молекул.
 Рис. 11.1-1. Схема строения ДНК
Рис. 11.1-1. Схема строения ДНК
|
А. Жизненно важные признаки, без которых бактериальная клетка не может существовать, закодированы в нуклеоиде (см. раздел 4.2).
Б. Не жизненно важные признаки закодированы у бактерий во внехромосомных факторах наследственности : плазмидах, транспозонах, IS-последовательностях и умеренных бактериофагах. Без этих признаков бактериальная клетка может существовать в обычных условиях, но дополнительные признаки, закодированные во внехромосомных факторах наследственности, придают ряду клеток популяции дополнительные возможности выживания при изменении условий внешней среды и, за счет повышения гетерогенности популяции, усиливают адаптационные возможности последней. Например, признак устойчивости к пенициллину в обычных условиях не востребован и не влияет на жизнеспособность бактериальной клетки, но при появлении этого антибиотика во внешней среде выжить могут только те бактерии, которые имеют плазмиду, кодирующую соответствующий признак. А выжить в присутствии пенициллина смогут только те бактериальные популяции, в составе которых есть клетки, несущие соответствующую плазмиду.
1. По способности к саморепликации внехромосомные факторы наследственности делятся на две группы: автономные и неавтономные.
а. Автономными внехромосомными факторами наследственности у бактерий являются плазмиды. Это значит, что они способны реплицироваться самостоятельно, обладая собственным соответствующим опероном.
б. Неавтономными внехромосомными факторами наследственности у бактерий являются транспозоны, IS-последовательности и умеренные фаги. Не обладая собственным опероном, обеспечивающим репликацию, они могут реплицироваться только в молекуле ДНК, обладающей таким опероном – или в нуклеоиде или в плазмиде.
2. Все внехромосомные факторы наследственности у бактерий могут встраиваться в нуклеоид (и, соответственно, выходить из его состава). В зависимости от места встраивания в нуклеоид внехромосомные факторы наследственности также делятся на две группы.
а. Только в гомологичных участках могут встраиваться в нуклеоид плазмиды и умеренные бактериофаги. Это значит, что для конкретной плазмиды и для конкретного умеренного бактериофага есть только одно место в нуклеоиде, где эта конкретная плазмида или этот конкретный бактериофаг могут включиться в состав нуклеоида.
б. В любых участках могут встраиваться в нуклеоид транспозоны и IS-последовательности.
11.2. Плазмиды
Плазмиды – внехромосомные автономные факторы наследственности у бактерий.
А. Плазмиды, как и другие внехромосомные факторы наследственности бактерий, увеличивают генетическую гетерогенность бактериальной популяции. Это и можно рассматривать как их основную функцию, которые они выполняют вместе с транспозонами, IS-последовательностями и умеренными бактериофагами. Конкретно же плазмиды выделяют две функции : регуляторную и кодирующую.
1. Плазмиды могут нести те же гены, которые имеются в нуклеоиде. Если один из этих генов нуклеоида по какой-либо причине «замолчит», то «включиться идентичный гены плазмиды – нужный признак по-прежнему будет присутствовать в фенотипе бактериальной клетки. Эта функция плазмид называется регуляторной .
2. Кодирующая функция плазмид заключается в том, что в них могут находиться гены, отсутствующие в бактериальной хромосоме (т.е. нуклеоиде). Собственно эта функция плазмид совпадает с общей функцией всех внехромосомных факторов наследственности – повышать гетерогенность генотипа популяции.
Б. Как и большинство других внехромосомных факторов наследственности, плазмиды могут существовать в бактериальной клетке в двух состояниях .
1. Если плазмида находится вне нуклеоида, в цитоплазме, говорят о ее автономном состоянии (не путать с понятием «автономный фактор наследственности»).
2. Плазмида, включенная в состав нуклеоида, находится в интегрированном состоянии.
В. В зависимости от содержания tra-оперона (о котором подробнее будет сказано ниже) плазмиды также делятся на две группы.
1. Плазмиды, содержащие в своем состав tra-оперон, называются конъюгативными , потому что такие плазмиды могут обуславливать передачу генетического материала от донорской клетки к реципиентной в процессе конъюгации (см. раздел 13.1).
а. Tra-оперон детерминирует образование конъюгативных пилей .
б. Кроме этого tra-оперон детерминирует цепь событий, которая называется мобилизацией на перенос . При этом участок двухцепочечной ДНК сначала расплетается, затем одна его цепочка переходит через конъюгативную пилю в другую, реципиентную, бактериальную клетку, где уже достраивается комплиментарная вторая нить. Таким образом, донорская клетка при конъюгации не теряет генетический материал, а реципиентная – приобретает его (несмотря на кажущийся смысл определения конъюгации как «перенос генетического материала от донорской к реципиентной клетке с помощью конъюгативных пилей).
1 . Tra-оперон может мобилизовывать на перенос саму конъюгативную плазмиду (как это происходит в случае передачи при конъюгации плазмиды F + , см. раздел 11.3).
2 . Tra-оперон может мобилизовывать на перенос другую, неконъюгативную, плазмиду (как это происходит в случае передачи при конъюгации плазмиды RTF, см. раздел 11.4).
3 . Tra-оперон может мобилизовывать на перенос участок нуклеоида донорской клетки (как это происходит при конъюгации, обусловленной плазмидой Hfr, см. раздел 11.3).
2. Плазмиды, не несущие tra-оперона, называются, соответственно, неконъюгативными , т.к. не способны обуславливать процесс конъюгации.
Г. На две группы делятся плазмиды и по степени контроля над их репликацией со стороны нуклеоида.
1. Большие плазмиды (т.е. имеющие относительно большой молекулярный вес) находятся под строгим контролем со стороны нуклеоида над своей репликацией. Нуклеоид «позволяет» им, как правило, лишь одно «лишнее» деление. Соответственно в клетке такие плазмиды находятся в одной или двух копиях.
2. Малые плазмиды (т.е. имеющие относительно малый молекулярный вес) находятся под ослабленным контролем со стороны нуклеоида над своей репликацией и могут делиться значительно чаще его – поэтому в бактериальной клетке может одновременно находится до 30 копий таких плазмид.
Д. Наконец, плазмиды классифицируются на группы несовместимости . Дело в том, что родственные плазмиды не могут одновременно находиться в одной бактериальной клетке, т.к. если одна из них уже в ней присутствует, то другая проникнуть в нее уже не сможет (такое явление называется иммунитет к суперинфекции и касается также взаимоотношения вирусов с чувствительной клеткой). В результате родственные друг другу плазмиды, не совместимые в одной клетке, составляют одну группу несовместимости. В настоящее время таких групп несовместимости насчитывается среди плазмид более двадцати.
11.3. F-плазмиды
F-плазмиды представляют собой собственно tra-оперон, без каких-либо дополнительных генов. Эту плазмиду называют еще половым фактором или фактором фертильности (откуда и название) потому что наличие F-плазмиды превращает клетку в возможного донора наследственного материала при конъюгации (т.е. в «мужскую» клетку), соответственно клетка, лишенная F-плазмиды, является потенциальным реципиентом генетического материала при конъюгации («женской» клеткой). Реципиентная клетка, получив при конъюгации F-плазмиду, превращается тем самым из женской в мужскую (т.е. происходит «смена пола»).
А. F-плазмида, находящаяся в интегрированном состоянии, носит название Hfr-плазмиды (high frequency of recombination – высокая частота рекомбинации). Если донорская клетка при конъюгации содержит Hfr-фактор, то при конъюгации разрыв цепочки нуклеоида происходит в месте прикрепления этого фактора и в конъюгационный мостик начинает входить противоположенный от места нахождения Hfr-фактора конец ДНК. Так как конъюгационный мостик не стоек, контакт между клетками при конъюгации кратковременен и вся цепочка ДНК нуклеоида просто не успевает перейти в реципиентную клетку. Следовательно, сам Hfr-фактор при этом виде конъюгации не передается (не происходит смена пола), а внедрившийся в реципиентную клетку фрагмент нуклеоида с высокой частотой (т.к. конъюгация происходит, как правило, между клетками одного вида) рекомбинирует с хромосомой реципиентной клетки.
Б. F-плазмида , находясь в автономном состоянии, сама переходит при конъюгации в реципиентную клетку. При этом реципиентная клетка превращается из «женской» в «мужскую» (т.е. происходит смена пола), но внедрившийся в реципиентную клетку генетический материал, представленный плазмидным генами, редко рекомбинирует с бактериальной хромосомой, потому что имеет по отношению к хромосомному генетическому материалу сравнительно низкую степень гомологии.
В. Половой фактор может переходить из автономного состояния в интеграционное и из интеграционного – в автономное. Последний процесс может сопровождаться обменом прилегающих частей плазмиды и нуклеотида, в результате чего перешедшая в автономное состояние плазмида окажется рекомбинационной – будет содержать фрагмент нуклеоида, оставив в нем взамен него свой фрагмент. Такая плазмида обозначается как F’-плазмида . Если донорская клетка при конъюгации несет F’-плазмиду, то она переходит в реципиентную клетку, производя у последней «смену пола», но при этом и обуславливает высокую частоту рекомбинации (благодаря включенному фрагменту нуклеоида). Формирование различных состояний F-плазмиды, описанное выше иллюстрируется на Рис. 11-3-1.
11.4. R-плазмиды
R-плазмиды – это плазмиды, детерминирующие множественную лекарственную устойчивость (или резистентность, откуда и название) бактериальной клетки к антибактериальным веществам (прежде всего антибиотикам).
А. Состав R-плазмиды определяется наличием в нем двух основных оперонов. Поэтому эта плазмида может находиться в двух формах.
1. Если в состав R-плазмиды входит tra-оперон, то в этом случае R-плазмида является конъюгативной . Tra-оперон в составе этой плазмиды называется RTF-фактор (resistance transfer factor – фактор, передающий устойчивость). Гены, детерминирующие устойчивость к антибиотикам, формируют так называемый r-оперон (если быть более точным, то устойчивость к каждому антибиотику детерминирует отдельный r-оперон – т.е. в состав R-плазмиды входит несколько r-оперонов), который, в свою очередь, тоже может существовать как самостоятельная плазмида. Другими словами, конъюгативная R-плазмида состоит из двух плазмид: RTF-фактора и r-фактора (что можно понимать как включение меньшей r-плазмиды в состав большей RTF-плазмиды).
2. Неконъюгативная форма этой плазмиды состоит только из r-оперона(-ов).
Б. В состав r-оперона может входить множество генов.
1. Во-первых, это гены, детерминирующие синтез специфических ферментов .
а. Это могут быть ферменты, инактивирующие антибиотик .
б. Или ферменты, модифицирующие антибиотик . При этом модифицированный антибиотик (например, фосфорилированный) теряет свою антибактериальную активность.
в. Наконец, это могут быть ферменты, которые снижают проницаемость клеточной стенки к данному антибиотику, в результате чего он не может проникнуть внутрь бактериальной клетки и «добраться» до мишени своего действия (например, до рибосом, чтобы блокировать здесь синтез белка).
2. И во-вторых, r-оперона может содержать целый транспозон или его часть – IS-последовательность (см. ниже).
В. Из состава r-оперона становятся понятны основные механизмы устойчивости бактерий к антибиотикам , обусловленные наличием R-плазмид. Их три.
1. R-плазмида может детерминировать способность бактериальной клетки инактивировать антибиотик .
2. Или R-плазмида может детерминировать способность бактериальной клетки таким образом модифицировать антибиотик , что он в результате потеряет свою антибактериальную активность.
3. И, наконец, R-плазмида может детерминировать снижение проницаемости клеточной стенки для данного антибиотика и, таким образом, закрыть ему доступ к мишени его действия.
Г. R-плазмида не всегда передается посредством конъюгации.
1. У грамположительных бактерий R-плазмида чаще всего передается посредством трансдукции .
2. Посредством конъюгации R-плазмида чаще всего передается у грамотрицательных бактерий.
11.5. Бактериоциногенные плазмиды (на примере Col-плазмиды E.coli)
Бактериоциногенные плазмиды присущи практически всем видам бактерий. Они детерминируют синтез антибиотикоподобных веществ – бактериоцинов. Мы рассмотрим эти плазмиды на примере кишечной палочки. У нее бактериоциногенные плазмиды называются колициногенными (как и у других бактерий – по названию вида) или Col-плазмидами (наиболее многочисленная группа колициногенных плазмид), а бактериоцины – колицинами (по такому же принципу).
А. Состав Col-плазмиды определяется наличием в нем двух основных оперонов.
1. Эта плазмида содержит, во-первых, гены, детерминирующие синтез колицинов .
а. Колицины представляют собой белки .
б. Насчитывается более 25 типов колицинов.
в. Колицины не действуют на клетку, несущую колициногенную плазмиду идентичного типа.
г. Колицины убивают бактериальную клетку, но не лизируют ее, что предотвращает появление в среде обитания оставшихся в живых бактерий токсических продуктов клеточного распада.
2. И, во-вторых, Col-плазмида содержит tra-оперон . Соответственно она относится к конъюгативным плазмидам.
Б. Col-плазмида отличается от других плазмид рядом присущих только ей особенностей .
1. В отличие от других конъюгативных плазмид Col-плазмида редко интегрирует в нуклеоид. Хотя все конъюгативные плазмиды по определению способны это делать, иначе они не смогли бы мобилизовывать на перенос участок бактериальной хромосомы (см. раздел 11.3.А).
2. Col-плазмида обычно репрессирована , т.е. информация с нее не снимается. Т.е. признак, который она детерминирует, в обычных условиях клетке не нужен.
3. Когда же данный признак становится востребован и Col-плазмида дерепрессируется, то бактериальная клетка синтезирует колицины и погибает. Другими словами, Col-плазмида является потенциально летальной .
В. Значение Col-плазмиды можно рассматривать с двух точек зрения.
1. Биологическое значение Col-плазмиды заключается в том, что с ее помощью достигается разрежение популяции при недостатке питательного субстрата. Происходит это по следующей схеме (Рис. 11.5-1).
а. В популяции присутствуют клетки с различными типами Col-плазмиды (на рисунке помечены различным цветом). При истощении питательной среды Col-плазмида в одной из клеток дерепрессируется, эта клетка синтезирует соответствующий тип колицина и погибает.
б. Колицины действуют только на те клетки, которые не содержат Col-плазмиду, детерминирующую синтез колицинов именно этого типа. Соответственно клетки, содержащие Col-плазмиду, детерминирующую синтез колицинов данного типа остаются живыми.
в. Все же другие клетки популяции, несущие Col-плазмиды других типов, погибают, популяция разряжается («едоков» становится меньше) и, как результат, может продлить свою жизнь на обедненной среде.
2. Медицинское значение Col-плазмида плазмиды заключается в том, что с ее помощью кишечные палочки, в норме заселяющие кишечник человека, регулируют нормальный микробиоценоз – совокупность микроорганизмов, населяющих кишечных здорового человека, так как колицины действуют не только на клетки своего вида, но и на клетки других видов бактерий кишечной группы.
11.6. Транспозоны
Транспозоны, впрочем, как и IS-последовательности, часто называют «прыгающими генами». Вследствие своей способности, в отличие от плазмид и умеренных фагов, внедряться в молекулу ДНК (нуклеоид, плазмиду или умеренный фаг) в любых, а не только в гомологичных, участках, они могут менять место своей локализации в той молекуле ДНК, в которой они интегрированы. Кроме того, они могут «перескакивать» также и из одной молекулы ДНК в другую.
А. Транспозонам обычно дается следующее определение : это нуклеотидные последовательности (от 2 000 до 20 000 пар нуклеотидов), способные менять место своей локализации в молекуле ДНК и мигрировать из одной молекулы ДНК в другую.
Б. В бактериальной клетке они могут находиться в двух состояниях .
1. Транспозон может находиться в интегрированном состоянии, т.е. быть встроенным в репликон (нуклеоид или плазмиду) и, соответственно, реплицироваться в составе этого репликона.
2. Транспозон может находиться в бактериальной клетке и в автономном состоянии (т.е. вне репликона). В этом случае он замыкается в кольцо, но, будучи не в состоянии самореплицироваться, при делении клетки переходит только в одну из двух вновь образованных.
В. Транспозон состоит из трех основных частей.
1. Он содержит особые концевые структуры , которые отличают транспозон от других фрагментов ДНК, встречающихся в бактериальной клетке, и являющихся, таким образом, маркерами именно этого внехромосомного фактора наследственности.
2. Способность транспозона менять место своей локализации в молекуле ДНК и мигрировать из одной молекулы ДНК в другую детерминируют особые гены транспозиции (собственно, это и есть IS-последовательности).
3. Кроме этого, транспозон содержит гены, детерминирующие синтез белков , обуславливающих наличие у содержащей данный транспозон бактериальной клетки дополнительных признаков.
а. Часто такие гены детерминируют способность бактериальной клетки синтезировать тот или иной белковый токсин .
б. Не менее часто транспозон детерминирует устойчивость клетки к какому-либо антибиотику (именно одному, в отличие от R-плазмиды).
в. Реже транспозон детерминирует синтез белков, обуславливающих другие признаки .
11.7. IS-последовательности
IS-последовательности (insert sequences – вставные последовательности) входят в состав транспозона, обеспечивая его способность к транспозиции.
А. IS-последовательностям обычно дается следующее определение : это вставки нуклеотидных последовательностей (порядка 1 000 пар нуклеотидов), способных к транспозиции.
Б. IS-последовательности принципиально отличаются от транспозонов .
1. Во-первых, они содержат только гены транспозиции .
2. Во-вторых, они не обнаружены в свободном состоянии .
В. IS-последовательности выполняют три основные функции .
1. С их помощью осуществляется координация взаимодействия внехромосомных факторов наследственности между собой и с бактериальной хромосомой для обеспечения их рекомбинации.
2. Кроме этого они могут осуществлять регуляторную функцию (т.е. регулировать транскрипции генов путём их «включения/выключения»).
3. Наконец, IS-последовательности могут индуцировать мутации (инверсии, дупликации на протяжении 5-9 пар нуклеотидов).
11.8. Механизмы фаговой конверсии
Здесь, после изложения сведений о бактериофаге и о внехромосомных факторах наследственности у бактерий, можно обобщить сведения о механизмах фаговой конверсии (см. раздел 10.4.Б.2), когда в результате лизогенизации (т.е. встраивание в геном умеренного бактериофага) бактерия приобретает дополнительный признак.
А. Во-первых это явление может быть обусловлено дерепрессией гена профага , как об этом было сказано выше (см. раздел 10.4.Б.2).
Б. Во-вторых дополнительный признак может детерминироваться геном бактерии-донора, внесенным в геном бактерии-реципиента дефектным фагом при специализированной трансдукции (см. раздел 10.4.В.2).
В. И в-третьих, если умеренный бактериофаг интегрировался около поврежденного промотора одного из генов, то функцию этого неработающего промотора может «взять на себя» промотор профага, восстановив тем самым экспрессию «молчавшего» гена бактерии-реципиента.
Похожая информация.
тема: Генетика бактерий
1865 год Мендель установил существование генов. 1869 Фишер выделил ДНК. Через 80 лет доказано что носителем генов является ДНК, 1953 Крик, Уотсон – расшифрована структура ДНК.
Ген выполняет следующие основные функции :
Непрерывность наследования генетической информации благодаря механизму репликации ДНК
Управление структурами и функциями организма с помощью генетического кода
Благодаря мутации и генетическим рекомбинациям, которые происходят в гене осуществляется эволюция всех живых организмов.
Код триплетный → кодон состоит из 3 букв и кодируют одну аминокислоту
Код не перекрывающийся
Число бессмысленных кодонов очень маленькое (3 из 64)
Последовательность расположения кодов в гене определяет последовательность положения аминокислотных остатков в полипептидной цепи
Код универсален
Способность к самоудвоению с помощью механизма саморепликации
Самовыражение (экспрессия) с помощью регулируемого синтеза матричной РНК
Самообновление с помощью мутаций, рекомбинаций и самоподвижных элементов
Самоисправляемая (ревизия, репарация, супрессия)
Гены вирусов и эукариотов состоят из экзона (кодирующий) и интроны (не кодирующие). У вирусов в одном и том же фрагменте могут существовать 2 гена с разными рамками считывания. Ген не всегда строго ограниченный участок хромосомы, есть подвижные участки у бактерий. Ген требует регулирования (регуляторы, промотеры). Ген единственные носитель и хранитель жизни, а белок определяет форму и способ жизни.
Эволюция генетической системы шла в направлении кодон(триплет) → ген → оперон → геном вирусов и плазмид → хромосома прокариотов → хромосома эукариотов (ядро).
Объем генома у представителей различных живых организмов сильно отличается. Можно измерить в следующих единицах: молекулярная масса нуклеиновых кислот либо в количестве нуклеотидных пар либо в количестве генов. Все эти значения сопоставимы ген в среднем включает 1000 пар нуклеотидов (вируса гепатита В – 4 гена; ВИЧ – 9 генов)
Генотип – вся совокупность генов у данного вида организма. 10%-70% не кодирующие гены (повторяющиеся последовательности), они не относятся к генотипу и составляют геном.
Фенотип – внешние проявления генотипа в конкретных условиях внешней среды при изменении внешних условий меняется генотип, но генотип при этом сохраняется.
^ Особенности генетики бактерий.
Хромосомы бактерий располагаются свободно в цитоплазме, не ограничены мембранами, но во всех случаях ДНК бактерии связана с рецепторами на мембране.
Бактерии гаплоидны, содержание ДНК не постоянно, может достигать 2, 4, 6, 8 – хромосом (у других организмов оно постоянно и удваивается только перед делением).
Передача генетической информации идет не только по вертикале (материнская→дочерняя), но и по горизонтали (конъюгация, трансформация)
Помимо хромосомного генома имеется не хромосомный генетический материал, который называется плазмидным геномом (эписомы, внехромосомные факторы наследственности). Это наделяет клетку дополнительными биологическими свойствами.
Содержание ДНК у бактерий зависит от условий их роста или от времени клеточного цикла бактерии, которые осуществляется каждые 20-30 минут, поэтому и количество может соответствовать (4,6,8) и это сопровождается увеличением количества рибосом (этапы транскрипции, трансляции идут одновременно, возможность регулировать скорость размножения главное условие сохранения вида.
^ Особенности репликации.
Вегетативная репликация: обуславливает передачу информации по вертикали, контролируется хромосомными и плазмидными генами.
Конъюгативная репликации: перенос материала по горизонтали и контролируется только плазмидными генами, при этом происходит достройка нити ДНК комплиментарной нити от донора к реципиенту.
Репаративная репликация: механизм при котором устраняется из ДНК поврежденный участок
Стркуктурно-функциональной едициней является оперон – группа структурных генов связанных с особым геном оператором, он управляет всей группой структурных генов и идет как самостоятельная единица, находится под контролем гена модулятора. В хромосоме гены распределяется друг за другом контролируя разные процессы, но законченный результат можно получить выбирая не последовательно (как игра на пианино).
^ Хромосомная карта бактерий
ромосомы бактерий имеют кольцевую форму, гены располагаются линейно, их можно последовательно расположить. Локализация генов определяют в минутах их переноса, и хромосомная карта это 0-100 минут.
Определение локализации гена на хромосоме называется картированием, а их расположение хромосомной картой масштаб которой в минутах. В настоящее время есть карты: кишечной палочки.
^ Изучение организации генома бактерий.
Проводится с помощью ферментов – рестриктаз способные расщепить ДНК в специфических участках, которые они комплиментарны. В настоящее время известно более 100 рестриктаз. С помощью них можно получить рестрикционные фрагменты ДНК → рестрикционный анализ. Сравнение рестрикционных фрагментов и называется рестрикционным анализом, который может быть использован для идентификации. Делают копии цепей ДНК, которые имеют липкие концы с помощью которых фрагменты вновь могут образовывать кольца. Именно за счет липких концов можно получать между разными фрагментами ДНК – рекомбинантные ДНК. Если эти фрагменты получены с помощью одной рестриктазы они могут вступать во взаимодействия между собой.
Метод клонирования. Выделенный фрагмент ДНК с помощью рекомбинантных молекул вводится в самореплицирующую генетическую структуру – в плазмиду, вирус и дальше они выполняют роль вектора для клонирования. Их сшивают с фрагментом ДНК – геномом, который будет размножаться в составе плазмилы или в составе геном бактериальной клетки. Такие гибридные ДНК также можно выделить из клетки за счет рестрикции – вырезания. С помощью клонирования можно получать большое количество копий любого фрагмента ДНК, который можно метить радиоактивной меткой.
Метод сегвинирования. Используют для определения последовательности расположение ДНК в клонируемом фрагменте ДНК. Методы секвинирования и клонирования это методы помогающие изучить геномы в т. ч. геном человека (2004).
^ Плазмидный геном бактериальной клетки.
Плазмиды – фрагменты ДНК с небольшой молекулярной массой, несут от 40 до 50 генов. Они выполняют также регуляторную и структурную функцию. Плазмиды могут располагаться либо в цитоплазме, могут иметь кольцевую структуру. Могут находится в интегрированном состоянии хромосомы (эписомы).
Свойства плазмид:
Не обязательные генетические элементы бактерий (дополнительные).
Обладают саморепликацией и автономностью, независимостью от хромосомы клетки. ДНК бактерии им не управляет.
Склонны к трансмиссии как по вертикали, так и по горизонтали обеспечивая при этом гегетическую изменчивость бактерий.
F-фактор – кольцевая молекула. Ее гены кодируют образование половых ворсинок, размножение бактерий, скорость размножения с ней связывают конъюгацию, участвует в горизонтальной передаче генетического материала и передаются различные свойства: устойчивость к антибиотикам, лактозо положительность.
R-фактор – детерминирует продукцию фермента β-лактамызы → устойчивость к антибиотикам. В составе этой плазмиды может быть специальный tra-оперон (ген отвечающий за перенос) → плазмида легко передается.
Hly – плазмида связана с продукцией гемотоксина → более токсигенные бактерии.
Col-фактор отвечает за продукцию колицинов (антибиотикоподобные вещества) обеспечивающих преимущество бактерий перед другими.
Плазмиды био деградации: участвуют в расщепдлении веществ загрязняющих окружающей среды.
Плазмида умеренного фага - фаг который способен распознать, внедрится, в клетку, но вызвать лизис бактерии вызвать не может. Может покидать клетку, захватывать часть генетического материала клетки и внедряясь в другую клетку участвует в переносе генетического материала (трансдукция)
Плазмиды есть конъюгативные (способные к переносу, имеющие в своем составе ген переноса), неконъюгативные (не участвуют в рекомбинации). По совместимости есть несовместимые друг с другом, совместимые.
^ Транспазоны, IS-последовательности.
Относятся к дополнительным генетичесим элементам
Th-маленькие участки ДНК (прыгающие) - в составе могут быть Rгены. Могут находится как в составе ДНК, так и в составе плазмид. Странспазонами связны мутации бактерии поскольку они могут перемещаться и вызывать мутации типа делеции, инверсии, дупликации.. Транспазоны ограничены с двух сторон IS-последовательностями.
IS-фрагменты – маленькие фрагменты ДНК, повторяющиеся, не способны к репродукции в свободном состоянии не участвуют. Основные функции: регуляторные (способны включить - выключить ген). Координируют взаимодействие транспазонов плазмид, фагов как между собой так и с хромосомой клетки хозяина.
^ Изменчивость бактерий.
Модификационная: адаптивная реакция организмов в ответ на условия внешней среды. Могут изменять морфологические, культуральные, ферментативные свойства.
Генотипическая: затрагивает генотип клетки:
Мутационная – изменение первичной структуры ДНК, могут быть связаны с выпадением нуклеатида, делецией могут носить характер инверсии. Могут быть хромосомные, плазмидные. Могут быть спонтанные, индуцированные. Значение эволционные изменение, сопроваждается селекцией.
Комбинативная: трансформация – передача генетического материала в виде раствора ДНК донора к реципиенту, трансдукция – перенос генетического материала от донора к реципиенту с помощью умеренных фагов (неспецифическая, специфическая), конъюгация – передача генетического материала от донора имеющего F-фактор к реципиенту через половые ворсинки с образованием новых штаммов.
Особенности генетики вирусов .
Молекулярная масса геном вирусов 10 6 меньше чем масса эукариотической клетки.
Организация генетического аппарата такая же
Генов от нескольких единиц до десятков.
Принцип 1 ген – молекул РНК – 1 белок у вирусных ДНК нарушен и иРНК вирусов может направлять синтез 2 и более белков.
Двукратное считывание одной и той же и РНК, но с другого кодона.
Сдвиг рамки трансляции
Сплайсинг (вырез интронов)
Транскрипция с перекрывающихся областей нуклеиновой кислоты → размывается границы гена и понятие ген приобретает функциональное значение.
Модификационная . В основном для вирусов определяет клетка хозяина. Модификация затрагивает суперкапсид.
Генотипическая . Мутационная, то есть изменение в первичной структуре нуклеотидов.
Рекомбинативная . Происходит при одновременном заражении клетки хозяина двумя или более вирусами, происходит обмен генами → образуются рекомбинантные штаммы вирусов, которые содержат гены 2 и более штаммов.
^ Генетическая реактивация . Процесс при котором вирионы дополняют друг друга в следствии перераспределения генов во время их репликации. Это наблюдается у вирусов с фрагментарным геномом. При скрещивании таких вирусов происходит образование полноценных единиц.
Комплементация (дополнение). Не генетически й процесс при котором вирус снабжает своего партнера (как правило дефектного) недостающими компонентами белка, а не нуклеиновыми кислотами. Характерна для многих вирусов – аденовирусы могут культивироваться только в присутсвии SV 40 – вирус. Вирус гепатита В является помощником для δ - вируса (HDV).
^ Фенотипическое смешивание . Наблюдается при совместном культивировании двух вирусов наблюдаем, что геном одного вируса заключается в капсид другого вируса. Генотип при этом не меняется
^ Генная инженерия.
Биотехнология использование биологических объектов (клеток микроорганизмов, грибов, животных, людей) для получения полезных для человека продуктов, которые не могут быть получены другим путем. Основное направление это генная инженерия. Появилась с 1972 когда появилась первая работа по генной инженерии.
Объект генной инженерии: ген или группа генов.
Источники получения: вирусы, прокарилты
Цель: пересадка гена в другие, гетерогенные системы, экспрессия этого гена и т.о. получать полезные продукты (белки, фермены, гормоны, лекарственные препараты и другие БАВ)
Инструмент генной инженерии: ферменты рестриктазы с помощью которых можно получать фрагменты генома. Рестриктазы имеют липкие концы для сшивания различных генов. Если их нет используют лигазы .
Этапы генной инженерии:
выделение гена из клетки с помощью рестриктаз из генома клетки.
присоединение гена к вектору (переносчику) – плазмиду, ДНК, РНК втрусов, умеренные фаги, искусственные плазмиды. Основные требования к вектору – должен выполнять роль саморепликации. Этот этап сопроваждается образованием рекомбинантной ДНК (ген+вектор)
введение рекомбинантной ДНК в гетерогенную систему. В качестве этой системы выступает клетка прокариотов, эукариотов, соматическая.
экспрессия введенного гена, создаются условия что бы рекомбинантная молекула начала самореплицироваться и заставила клетку продуцировать вещество, которое кодирует перенесенный ген.
клонирование гена и выделение продукта, очитка продукта и выхода продукта