Что такое структурный ген. Понятие о гене. Классификация генов. Структурная организация генов прокариот и эукариот. Регуляция работы генов у эукариот
Фундаментальным понятием в генетике является представление о гене как единице наследственности. Ниже приводится два определения гена. Ген - это участок ДНК, коллинеарно кодирующий определённый белковый или нуклеиновый продукт. Ген - это фрагмент 2-цепочечной ДНК, несущей определённую генетическую информацию. У кишечной палочки имеется 4 тыс. генов, у дрожжей - 7 тыс. генов, а у дрозофилы и плоских червей -15-20 тысяч генов,У человека имеется приблизительно от 50 тысяч до 100 тысяч структурных генов, по данным на 1989 год около 5 тысяч генов были приблизительно охарактеризованы, а около 2 тысяч генов были нанесены на карты хромосом (картированы). 26 июня 2000 года в прессе было сделано сообщение о том, что учеными США, Англии, Японии и других стран, участвующими в программе «Геном человека», завершена основная часть работы (более 90%) по расшифровке генетического кода человека.
В ближайшие 2 года планируется уточнить и завершить работу по данной программе, которая имеет важное прикладное значение для медицины. В плане данной темы важно помнить, что ген занимает определённый участок (локус) в хромосоме, это участок ДНК, который может быть представлен десятками, сотнями или тысячами пар нуклеотидов. В настоящее время, с функционально-генетической точки зрения, гены классифицируют на 3 группы:
- 1. Структурные гены - кодируют структуру синтезируемых клеткой белков (структурных белков, белков-ферментов и др.), а также кодируют последовательности нуклеотидов в молекулах т-РНК и р-РНК.
- 2. Регуляторные (функциональные) гены - контролируют и направляют работу структурных генов.
- 3. Гены-модуляторы. К ним относятся гены-ингибиторы (или супрессоры), которые подавляют функции других генов, гены-интенсификаторы, которые усиливают функции других генов и др. Экзонно-интронная структура генов.
В 70-х годах XX века было обнаружено, что структурные гены эукариот содержат экзоны (участки ДНК, несущие генетическую информацию и отвечающие за синтез определенных участков белков) и интроны (участки ДНК, которые не несут генетической информации, относящейся к синтезу белка, кодируемого данным геном). Интроны ещё называют вставками, расположенными между экзонами. Таким образом, принципиальным отличием генов эукариот от генов прокариот является то, что их структурные гены имеют разорванную, прерывистую структуру. Однако исключение составляют гены, кодирующие гистоны и интерфероны, они не содержат интронов. Дальнейшие исследования показали, что большинство генов эукариот имеют экзон-интронную организацию. Длина интронов варьирует в очень широких пределах: от 100 до 10000 нуклеотидов и более, нередко их суммарная длина больше длины экзонов. Количество интронов и экзонов в разных генах варьирует. Один из самых коротких - ген бета-глобина, состоящий из 1100 пар нуклеотидов (пн), содержит 3 экзона (90, 222, 126 пн) и 2 интрона (116, 646 пн). Примером протяженного гена служит ген дистрофина, имеющий 2,6 млн пн и более 2000 экзонов.
Представление, что интроны - нефункциональная часть гена, - неверно. И хотя детально их биологическая роль не выяснена, существует ряд гипотез о значении интронов:
1) Строение генов из участков выгодно для процессов генетической рекомбинации, перетасовки генов. Чем дальше в хромосоме расположены фрагменты генетического материала, тем выше вероятность рекомбинации. Именно поэтому и выгодны вставки-интроны. Нуклеотидная последовательность интронов менее консервативна, чем у экзонов, она подвергается быстрым изменениям в эволюции.
Перетасовка частей генов может быть использована для разных целей: а) это путь к образованию новых генов; б) это способ нейтрализации вредных мутаций.
2) Предполагается регуляторная роль интронов в экспрессии (работе) генов. Интроны могут содержать энхансеры. Они могут кодировать особый фермент, который участвует в сплайсинге м-РНК (смотри следующий вопрос). Заканчивая разговор о гене, необходимо отметить ещё одно обстоятельство. У эукариот гены разделены между собой протяженными участками ДНК, которые были названы спейсерами, или разделителями. Накапливается всё более данных, что именно в спейсерах располагаются те сегменты ДНК, которым принадлежит решающая роль в регуляции работы генов (в регуляции транскрипции). Регуляиия биосинтеза белка у прокариот (на примере работы лактозного оперона кишечной папочки).
Все клетки любого организма имеют полный набор свойственных данному организму генов. Вместе с тем известно, что клетки разных тканей и органов отличаются по набору имеющихся в них белков. Располагая полной генетической информацией, каждая клетка на определенном этапе развития использует лишь ту её часть, которая необходима в настоящий момент, транскрибируются («работают») только те гены, продукты которых нужны клетке в данный момент для выполнения её функций. Следовательно, клетка должна обладать механизмами, определяющими какие гены и в какой последовательности должны транскрибироваться. Наиболее полно регуляция генной активности изучена на примерах синтеза белков-ферментов у микроорганизмов.
Теория регуляции биосинтеза белка у прокариот разработана в 50-х годах XX века французскими учеными Ф. Жакобом и Ж. Моно. Они разработали концепцию опреона и выяснили основные принципы регуляции биосинтеза белка у прокариот. Согласно теории Ф. Жакоба и Ж. Моно, гены функционально неодинаковы: выделяют группу структурных генов (они кодируют структуру синтезируемых клеткой попипептидов, белков, р-РНК, т-РНК) и группу регуляторных генов (они управляют работой структурных генов обычно с помощью присоединения к ним различных белковых факторов).
Единицей генетической регуляции является оперон, который представляет собой совокупность расположенных е линейной последовательности регуляторных и одного или нескольких структурных генов. Гены одного оперона расположены в хромосоме прокариот рядом и кодируют ферменты, осуществляющие последовательные реакции синтеза или расщепления. Эти гены находятся под общим регуляторным контролем и могут включаться и выключаться координированно. Одним из наиболее наглядных и хорошо изученных примеров является лактозный оперон кишечной палочки (Escherichia coli) - группа генов, контролирующая синтез ферментов, осуществляющих катаболизм молочного сахара - лактозы. Буквально через несколько минут после добавления в питательную среду для кишечной палочки лактозы, бактерии начинают вырабатывать 3 фермента: галактозидпермеазу, бетагалактозидазу и галактоэидтрансацетилазу. Как только ресурсы лактозы в среде исчерпываются, синтез ферментов сразу же прекращается.
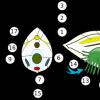
Строение лактозного оперона кишечной палочки:
- 1. Начинается оперон с участка А - он предназначен для присоединения белка-активатора (синий круглешок), в свою очередь необходимого для присоединения к следующему участку фермента (РНК-полимеразы).
- 2. Следующий участок П (промотор) - место прикрепления фермента РНК-полимеразы (зеленый треугольник), это участок начала транскрипции.
- 3. За промотором следует О (оператор) - он играет важную роль в транскрипции генов оперона, т.к. с ним может прикрепляться регуляторный белок-репрессор (красн 2 треугольника)
- 4. За оператором следуют структурные гены (z, у, а), которые кодируют построение 3-х упомянутых ранее белков-ферментов.
- 5. Заканчивается оперон Т (терминатором) - участком, прекращающим продвижение РНК-полимеразы и транскрипции оперона.
- 6. Основная регуляция работы структурных генов осуществляется регуляторным белком (красн 2 треугольн) который кодируется Р (геном-регулятором), который не входит в состав оперона, а лежит поблизости в другом месте хромосомы.
Работа лактозного оперона Регуляторный белок-репрессор в незначительном количестве синтезируется в клетке постоянно. Этот белок обладает сродством к последовательности нуклеотидов в области оператора, а также сродством к лактозе. Репрессия: В отсутствие лактозы регуляторный белок связывается с участком-оператором (О) и препятствует продвижению по ДНК РНК-полимеразы: не синтезируется м-РНК, не синтезируются и белки-ферменты. Индукция: После добавления в среду лактозы, регуляторный белок связывается с ней быстрее, чем с участком-оператором, который остаётся свободным и не препятствует продвижению РНК-полимеразы. Идёт транскрипция и трансляция. Синтезирующие белки-ферменты расщепляют лактозу. После того, как вся лактоза будет израсходована, нечем будет связывать регуляторный белок и он снова окажется с О (оператором), прекратив транскрипцию оперона.
Другой известный тип индукции - позитивная индукция. Она свойственна другому оперону кишечной палочки, кодирующему ферменты катаболизма другого сахара - арабинозы. Этот оперон структурно очень похож на предыдущий. Разница в регуляции состоит в том, что добавленная в среду арабиноза взаимодействует с белком-репрессороми, освобождая операторный участок, одновременно превращает белок-репрессор в белок-активатор, способствующий. присоединению РНК-полимеразы к промотору. В этих условиях транскрипции имеет место. Как только запасы арабинозы в среде исчерпываются, синтезирующийся белок-репрессор опять связывается с оператором, выключая транскрипцию.
Кроме индукции, известны также 2 типа (негативный и позитивный) регуляции по принципу репрессии. Если при негативной индукции эффектор (индуктор) препятствует присоединению белка-репрессора к оператору, то при негативной репрессии, наоборот, эффектор придаёт регуляторному белку способность присоединяться к оператору. Если в первом случае соединение эффектора с белком-регулятором разрешало транскрипцию, то во втором оно запрещает её. Примером негативной репрессии может служить хорошо изученный триптофановый оперон кишечной палочки.
В его состав входят пять структурных генов, обеспечивающих синтез аминокислоты триптофана, оператор и два промотора. Белок-регулятор синтезируется вне триптофонового оперона. Пока клетка успевает расходовать весь синтезирующийся триптофан, оперон работает, синтез триптофана продолжается. Если же в клетке появляется избыток триптофана, он соединяется с регуляторным белком и изменяет его таким образом, что этот белок приобретает сродство с оператором. Измененный белок-регулятор взаимодействует с оператором и препятствует транскрипции структурных генов вследствии чего синтез триптофана прекращается. При позитивной репрессии эффектор лишает регуляторный белок способности связываться с оператором, обуславливая таким образом, транскрипцию структуоных генов.
Описанные типы регуляции характеризуют механизмы регуляции отдельных оперонов, практически не касаясь регуляции экспрессии генома в целом, в то время как совершенно очевидно, что регуляция разных оперонов должна носить согласованный характер. Такой согласованный характер работы разных оперонов и генов получил у вирусов и фагов название каскадной регуляции. Согласно принципу каскадной регуляции, сначала происходит транскрипция «предранних», затем «ранних» и наконец «поздних» генов, в зависимости от того, какие белки требуются на разных стадиях вирусной (фаговой) инфекции.
Конечно, принцип каскадной регуляции у фагов относится к наиболее простым. У более сложно организованных организмов для осуществления большого количества функций, происходящих одновременно или с определённой последовательностью, необходима согласованная работа многих генов и оперонов, Особенно это касается эукариотов, отличающихся не только более сложной организацией генома, но и многими другими особенностями механизмов регуляции генной активности.
По принципам регуляции гены эукариотов можно условно разделить на 3 группы:
- 1) функционирующие во всех клетках организма;
- 2) функционирующие в тканях только одного типа;
- 3) обеспечивающие выполнение специализированными клетками конкретных функций.
Кроме того, у эукариотов известно одновременное групповое выключение генной активности, осуществляемое гистонами - основными белками, входящими в состав хромосом. Ещё одним существенным отличием транскрипции у эукариотов является то, что многие м-РНК длительное время сохраняются в клетке в виде особых частиц -информосом, в то время как м-РНК прокариотов практически ещё в процессе транскрипции поступают в рибосомы, транслируются, после чего быстро разрушаются.
Вместе с тем, имеется много данных, указывающих, что транскрипция у эукариотов осуществляется с участков, подобных оперонам прокариотов и состоящих из регуляторных и структурных генов. Отличительной особенностью оперонов эукариотов является то, что почти всегда они содержат только структурный ген, а гены, контролирующие различные этапы определённой цепи метаболических превращений разбросаны по хромосоме и даже по разным хромосомам.
Другой отличительной чертой оперонов эукариотов является то, что они состоят из значащих (экзонов) и незначащих (интронов) участов. чередующихся друг с другом. При транскрипции считываются как экзоны, так и интроны, а образующийся при этом предшественник информационной РНК (про-мРНК) затем претерпевает созревание (процессинг), в результате которого происходит вырезание интронов и образование собственно м-РНК (сплайсинг).
У эукариотов известны и другие типы регуляции активности генов, такие как эффект положения или дозовая компенсация. В первом случае речь идёт об изменении генной активности е зависимости от конкретного окружения: перемещение гена из одного места хромосомы в другое может приводить к изменению активности как этого гена, так и близлежащих. Во втором случае, нехватка одной дозы какого-либо гена (в первую очередь это относится к генам, локализованным в половых хромосомах гетерогаметного пола, когда одна из гомологичных половых хромосом либо генетически инертна, либо полностью отсутствует) фенотипически не проявляется за счет компенсаторного увеличения активности оставшегося гена, В целом же, регуляция активности генов у эукариотов изучена недостаточно.
Ген (др.-греч. γένος — род) — структурная и функциональная единица наследственности живых организмов. Ген представляет собой последовательность ДНК, задающую последовательность определённого полипептида либо функциональной РНК. Гены (точнее, аллели генов) определяют наследственные признаки организмов, передающиеся от родителей потомству при размножении. При этом некоторые органеллы (митохондрии, пластиды) имеют собственную, определяющую их признаки, ДНК, не входящую в геном организма.
В настоящее время, в молекулярной биологии установлено, что гены — это участки ДНК, несущие какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК. Эти и другие функциональные молекулы определяют развитие, рост и функционирование организма.
В то же время, каждый ген характеризуется рядом специфических регуляторных последовательностей ДНК, таких как промоторы, которые принимают непосредственное участие в регулировании проявления гена. Регуляторные последовательности могут находиться как в непосредственной близости от открытой рамки считывания, кодирующей белок, или начала последовательности РНК, как в случае с промоторами (так называемые cis-регуляторные элементы, англ. cis-regulatory elements), так и на расстоянии многих миллионов пар оснований (нуклеотидов), как в случае с энхансерами, инсуляторами и супрессорами (иногда классифицируемые как trans-регуляторные элементы, англ. trans-regulatory elements). Таким образом, понятие гена не ограничено только кодирующим участком ДНК, а представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности.
Изначально термин ген появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулы могут являться носителями наследственной информации. Большинство исследователей считали, что такими носителями могут быть только белки, так как их строение (20 аминокислот) позволяет создать больше вариантов, чем строение ДНК, которое составлено всего из четырёх видов нуклеотидов. Позже было экспериментально доказано, что именно ДНК включает в себя наследственную информацию, что было выражено в виде центральной догмы молекулярной биологии.
Гены могут подвергаться мутациям — случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК, которые, в свою очередь, могут иметь результатом общее или локальное изменённое или анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание, или летальными на эмбриональном уровне. Однако далеко не все изменения последовательности нуклеотидов приводят к изменению структуры белка (благодаря эффекту вырожденности генетического кода) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуется однонуклеотидными полиморфизмами и вариациями числа копий (англ. copy number variations), такими как делеции и дупликации, которые составляют около 1 % всей нуклеотидной последовательности человека. Однонуклеотидные полиморфизмы, в частности, определяют различные аллели одного гена.
Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие в себя азотистые основания: аденин(А) или тимин(Т) или цитозин(Ц) или гуанин(Г), пятиатомный сахар-пентозу-дезоксирибозу, по имени которой и получила название сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов.
Свойства гена.
дискретность — несмешиваемость генов;
стабильность — способность сохранять структуру;
лабильность — способность многократно мутировать;
множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
аллельность — в генотипе диплоидных организмов только две формы гена;
специфичность — каждый ген кодирует свой признак;
плейотропия — множественный эффект гена;
экспрессивность — степень выраженности гена в признаке;
пенетрантность — частота проявления гена в фенотипе;
амплификация — увеличение количества копий гена.
Строение гена.
Согласно современным представлениям, ген, кодирующий синтез определенного белка, у эукариот состоит из нескольких обязательных элементов. Прежде всего это обширная регуляторная зона, оказывающая сильное влияние на активность гена в той или иной ткани организма на определенной стадии его индивидуального развития. Далее расположен непосредственно примыкающий к кодирующим элементам гена промотор - последовательность ДНК длиной до 80-100 пар нуклеотидов, ответственная за связывание РНК-полимеразы, осуществляющей транскрипцию данного гена. Вслед за промотором лежит структурная часть гена, заключающая в себе информацию о первичной структуре соответствующего белка. Эта область для большинства генов эукариот существенно короче регуляторной зоны, однако ее длина может измеряться тысячами пар нуклеотидов.
Важная особенность эукариотических генов - их прерывность. Это значит, что область гена, кодирующая белок, состоит из нуклеотидных последовательностей двух типов. Одни - экзоны - это участки ДНК, которые несут информацию и строении белка и входят в состав соответствующих РНК и белка. Другие - интроны - не кодируют структуру белка и в состав зрелой молекулы и-РНК не входят, хотя и транскрибируются. Процесс вырезания интронов - «ненужных» участков молекулы РНК и сращивания экзонов при образовании и-РНК осуществляется специальными ферментами и получил название Сплайсинг (сшивание, сращивание). Экзоны обычно соединяются вместе в том же порядке, в котором они распологаются в ДНК. Однако не абсолютно все гены эукариот прерывисты. Иначе говоря, у некоторых генов, подобно бактериальным, наблюдается полное соответствие нуклеотидов последовательности первичной структуре кодируемых ими белков. Таким образом, ген эукариот во многом похож на оперон прокариот, хотя и отличается от него более сложной и протяженной регуляторной зоной, а также тем, что он кодирует обычно только один белок, а не несколько, как оперон у бактерии.
8.1. Ген как дискретная единица наследственности
Одним из фундаментальных понятий генетики на всех этапах ее развития было понятие единицы наследственности. В 1865 году основоположник генетики (науки о наследственности и изменчивости) Г. Мендель на основании результатов своих опытов на горохе пришел к заключению, что наследственный материал дискретен, т.е. представлен отдельными единицами наследственности. Единицы наследственности, которые отвечают за развитие отдельных признаков, Г.Мендель назвал «задатками». Мендель утверждал, что в организме по любому признаку имеется пара аллельных задатков (по одному от каждого из родителей), которые между собой не взаимодействуют, не смешиваются и не изменяются. Поэтому, при половом размножении организмов в гаметы попадает лишь один из наследственных задатков в «чистом» неизменном виде.
Позже предположения Г.Менделя о единицах наследственности получили полное цитологическое подтверждение. В 1909 году датский генетик В. Иогансен назвал «наследственные задатки» Менделя генами.
В рамках классической генетики ген рассматривается как функционально неделимая единица наследственного материала, определяющая формирование какого-либо элементарного признака.
Различные варианты состояния определенного гена, возникшие в результате изменений (мутаций), получили название «аллели» (аллельные гены). Количество аллелей гена в популяции может быть значительным, но у конкретного организма число аллелей определенного гена всегда равно двум – по числу гомологичных хромосом. Если в популяции количество аллелей какого-либо гена больше двух, то такое явление получило название «множественного аллелизма».
Гены характеризуются двумя противоположными по биологическому значению свойствами: высокой стабильностью своей структурной организации и способностью к наследственным изменениям (мутациям). Благодаря этим уникальным свойствам обеспечивается: с одной стороны – устойчивость биологических систем (неизменяемость в ряде поколений), а с другой – процесс их исторического развития, формирования адаптаций к условиям окружающей среды, т.е. эволюция.
8.2. Ген как единица генетической информации. Генетический код.
Еще Аристотель более 2500 лет тому высказал предположение о том, что гаметы – это отнюдь не миниатюрные варианты будущего организма, а структуры, содержащие информацию о развитии эмбрионов (хотя он признавал только исключительно важность яйцеклетки в ущерб сперматозоиду). Однако развитие этой идеи в современных исследованиях стало возможным лишь после 1953 года, когда Дж. Уотсон и Ф. Крик разработали трехмерную модель строения ДНК и тем самым создали научные предпосылки для раскрытия молекулярных основ наследственной информации. С этого времени началась эра современной молекулярной генетики.
Развитие молекулярной генетики привело к раскрытию химической природы генетической (наследственной) информации и наполнило конкретным смыслом представление о гене как единици генетической информации.
Генетическая информация – информация о признаках и свойствах живых организмов, заложенная в наследственных структурах ДНК, которая реализуется в онтогенезе через синтез белка. Наследственную информацию, как программу развития организма, каждое новое поколение получает от предков в виде совокупности генов генома. Единицей наследственной информации является ген, который представляет собой функционально неделимый участок ДНК со специфической последовательностью нуклеотидов, определяющей последовательность аминокислот определенного полипептида или нуклеотидов РНК.
Наследственная информация о первичной структуре белка записана в ДНК с помощью генетического кода.
Генетический код – система записи генетической информации в молекуле ДНК (РНК) в виде определенной последовательности нуклеотидов. Этот код служит ключом для перевода последовательности нуклеотидов в и-РНК в последовательность аминокислот полипептидной цепи при ее синтезе.
Свойства генетического кода:
1. Триплетность – каждая аминокислота кодируется последовательностью из трех нуклеотидов (триплетом или кодоном)
2. Вырожденность – большинство аминокислот шифруется более чем одним кодоном (от 2 до 6). В ДНК или РНК имеется 4 различных нуклеотида, которые теоретически могут образовывать 64 разных триплета (4 3 = 64) для кодирования 20 аминокислот, входящих в состав белков. Этим и объясняется вырожденность генетического кода.
3. Неперекрываемость – один и тот же нуклеотид не может входить одновременно в состав двух соседних триплетов.
4. Специфичность (однозначность) – каждый триплет кодирует только одну аминокислоту.
5. Код не имеет знаков препинания. Считывание информации с и-РНК при синтезе белка всегда идет в направлении 5 , -- 3 , в соответствии с последовательностью кодонов иРНК. Если произойдет выпадение одного нуклеотида, то при считывание его место займет ближайший нуклеотид из соседнего кода, из-за чего изменится аминокислотный состав в молекуле белка.
6. Код универсален для всех живых организмов и вирусов: одинаковые триплеты кодируют одинаковые аминокислоты.
Универсальность генетического кода свидетельствует о единстве происхождения всех живых организмов
Однако, универсальность генетического кода не является абсолютной. В митохондриях ряд кодонов имеет другой смысл. Поэтому иногда говорят о квазиуниверсальности генетического кода. Особенности генетического кода митохондрий свидетельствует о возможности его эволюционирования в процессе исторического развития живой природы.
Среди триплетов универсального генетического кода три кодона не кодируют аминокислоты и определяют момент окончания синтеза данной полипептидной молекулы. Это так называемые «nonsens» кодоны (стоп-кодоны или терминаторы). К ним относятся: в ДНК – АТТ, АЦТ, АТЦ; в РНК – УАА, УГА, УАГ.
Соответствие нуклеотидов в молекуле ДНК порядку аминокислот в молекуле полипептида получило название коллинеарности. Экспериментальное подтверждение коллинеарности сыграло решающую роль в расшифровке механизма реализации наследственной информации.
Значение кодонов генетического кода приведены в таблице 8.1.
Табл.8.1. Генетический код (кодоны иРНК для аминокислот)
С помощью этой таблицы по кодонам иРНК можно определить аминокислоты. Первый и третий нуклеотиды берут из вертикальных столбиков, расположенных справа и слева, а второй – с горизонтального. В месте перессичения условных линий содержится информация про соответствующую аминокислоту. Отметим, что в таблице приводится триплеты и-РНК, а не ДНК.
Структурно - функциональная организация гена
Молекулярная биология гена
Современное представление о строении и функции гена формировалось в русле нового направления, которое Дж.Уотсон назвал молекулярной биологией гена (1978)
Важным этапом в изучении структурно – функциональной организации гена были работы С. Бензера в конце 1950-хх годов. Они доказали, что ген представляет собой нуклеотидную последовательность, которая может изменятся в результате рекомбинаций и мутаций. Единицу рекомбинации С.Бензер назвал реконом, а еденицу мутации – мутоном. Экспериментально установлено, что мутон и рекон соответствуют одной паре нуклеотидов. Единицу генетической функции С. Бензер назвал цистроном.
В последние годы стало известно, что ген имеет сложное внутренее строение, а отдельные его части обладают разными функциями. В гене можно выделить последовательность нуклеотидов гена, которая определяет строение полипептида. Эта последовательность называется цистроном.
Цистрон – это последовательность нуклеотидов ДНК, которая определяет отдельную генетическую функцию полипептидной цепи. Ген может быть представлен одним или несколькими цистронами. Сложные гены содержащие в себе несколько цистронов называются полицистронными .
Дальнейшее развитие теории гена связано с выявлением различий в организации генетического материала у организмов далеких друг от друга в таксономическом отношении, которыми являются про- и эукариоты.
Структура генов прокариот
У прокариот, типичными представителями которых являются бактерии, большинство генов представлены непрерывными информативными участками ДНК, вся информация которых используется при синтезе полипептида. У бактерий гены занимают 80-90% ДНК. Главная особенность генов прокариот – это их объединение в группы или опероны.
Оперон – это группа следующих подряд структурных генов, находящихся под контролем одного регуляторного участка ДНК. Все сцепленые гены оперона кодируют ферменты одного метаболического пути (например, расщепление лактозы). Такая общая молекула иРНК называется полицистронной. Только некоторые гены прокариот транскрибируются индивидуально. Их РНК называется моноцистронной.
Организация по типу оперона позволяет бактериям быстро переключать метаболизм с одного субстрата на другой. Бактерии не синтезируют ферменты определенного метаболического пути в отсутствии необходимого субстрата, но способны начать их синтезировать при появлении субстрата.
Структура генов эукариот
Большинство генов эукариот (в отличии от генов прокариот) имеют характерную особенность: содержат не только кодирующие структуру полипептида участки – экзоны, но и некодирующие – интроны. Интроны и экзоны чередуются между собой, что придает гену прерывистую (мозаичную) структуру. Количество интронов в генах варьиирует от 2-х до десятков. Роль интронов до конца неясна. Полагают, что они учавствуют в процессах рекомбинации генетического материала, а также в процессах регуляции экспресии (реализации генетической информации) гена.
Благодаря экзонно – интронной организации генов создаются предпосылки для альтернативного сплайсинга. Альтернативний сплайсинг- процесс «вырезания» разных интронов из первичного РНК-транскрипта в результате чего на основе одного гена могут синтезироватся разные белки. Явление альтернативного сплайсинга имеет место у млекопитающих при синтезе различних антител на основе иммуноглобулиновых генов.
Дальнейшие исследование тонкой структуры генетического материала еще больше осложнило четкость определения понятия «ген». В геноме эукариот были обнаружены обширные регуляторные области имеющие различные участки, которые могут располагатся за пределами едениц трансскрипции на расстоянии в десятки тысяч пар нуклеотидов. Структуру эукариотического гена, включающую транскрибируемые и регуляторные области, можно представить следующие образом.
Рис 8.1. Структура эукариотического гена
1 – энхансеры; 2 – сайленсеры; 3 – промотор; 4 – экзоны; 5 – интроны; 6 – участки экзонов, кодирующие нетранслируемые области.
Промотор – участок ДНК для связывания с РНК – полимеразой и образование комплекса ДНК-РНК полимеразы для запуска синтеза РНК.
Энхансеры - усилители транскрипции.
Сайленсеры – ослабители транскрипции.
В настоящее время ген (цистрон) рассматривается как функционально неделимая единица наследственного мастерства, определяющая развитие какого – либо признака или свойства организма. С позиции молекулярной генетики ген представляет собой участок ДНК (у некоторых вирусов РНК), который несет информацию о первичной структуре полипептида, молекулы транспортной и рибосомальной РНК.
В диплоидных клетках человека примерно 32000 пар генов. Большинство генов в каждой клетке «молчит». Набор активных генов зависит от типа ткани, периода развития организма, полученных внешних или внутренних сигналов. Можно сказать, что в каждой клетке «звучит» свой акорд генов, определяя спектр синтезируемых РНК, белков и, соответственно, свойства клетки.
Структура генов вирусов
Вирусы имеют структуру гена, отражающую генетическую структуру клетки - хозяина. Так, гены бактериофагов собраны в опероны и не имеют интронов, а вирусы эукариот имеют интроны.
Характерная особенность вирусных геномов – это явление «перекрывающихся» генов («ген в гене»). В «перекрывающихся» генах каждый нуклеотид принадлежит одному кодону, но имеются разные рамки считывание генетической информации с одной и той же нуклеотидной последовательности. Так, у фага φ Х 174 имеется участок молекулы ДНК, который входит в состав сразу трех генов. Но соответствующие этим генам последовательности нуклеотидов прочитывается каждая в своей системе отсчета. Поэтому нельзя говорить о «перекрывании» кода.
Такая организация генетического материала («ген в гене») расширяет информационные возможности сравнительно небольшого по величине генома вирусов. Функционирование генетического материала вирусов происходит по-разному в зависимости от структуры вируса, но всегда с помощью ферментной системы клетки хозяина. Различные способы организации генов у вирусов, про- и эукариотов представлены на рис 8.2.

Функционально – генетическая классификация генов
Существует несколько классификаций генов. Так, например, выделяют аллельные и неаллельные гены, летальные и полулетальные, гены «домашнего хозяйства», «гены роскоши» и т.д.
Гены «домашнего хозяйства» - набор активных генов, необходимых для функционирования всех клеток организма независимо от типа ткани, периода развития организма. Эти гены кодируют ферменты транскрипции, синтеза АТФ, репликации, репарации ДНК и др.
Гены «роскоши» имеют избирательную активность. Их функционирование специфично и зависит от типа ткани, периода развития организма, полученных внешних или внутренних сигналов.
Исходя из современных представлений о гене как функционально неделимой единице наследственного материала и системной организации генотипа все гены принципиально можно разделить на две группы: структурные и регуляторные.
Регуляторные гены – кодируют синтез специфических белков, влияющих на функционирование структурных генов таким образом, что в клетках разной тканевой принадлежности синтезируются необходимые белки и в необходимых количествах.
Структурными называются гены, которые несут информацию о первичной структуре белка, рРНК или тРНК. Гены, кодирующие белки, несут информацию о последовательности аминокислот определенных полипептидов. С этих участков ДНК транскребируется иРНК, которая служит матрицей для синтеза первичной структуры белка.
Гены рРНК (выделяют 4 разновидности) содержат информацию о последовательности нуклеотидов рибосомальных РНК и обуславливают их синтез.
Гены тРНК (более 30 разновидностей) несут информацию о строении транспортных РНК.
Структурные гены , функционирование которых тесно связано со специфическими последовательностями в молекуле ДНК, называемыми регуляторными участками, подразделяются на:
· независимые гены;
· повторяющиеся гены;
· кластеры генов.
Независимые гены – это гены, транскрипция которых не связана с транскрипцией других генов в рамках транскрипционной еденицы. Их активность может регулироваться экзогенными веществами, например, гормонами.
Повторяющиеся гены присутствуют в хромосоме в виде повторов одного гена. Ген рибосомной 5-S-РНК повторяется много сотен раз, причем повторы располагаются тандемом, т. е. следуя вплотную друг за другом без промежутков.
Кластеры генов – это локализованные в определенных участках (локусах) хромосомы группы разных структурных генов с родственными функциями. Кластеры тоже часто присутствуют в хромосоме в виде повторов. Например, кластер гистоновых генов повторяется в геноме человека 10-20 раз, обазуя тандемную групу повторов.(рис. 8.3.)

Рис.8.3. Кластер гистоновых генов
За редким исключением кластеры транскрибируются как одно целое – в виде одной длинной пре-мРнк. Так пре-мРНК кластера гистоновых генов содержит информацию про все пять гистоновых белков. Это ускоряет синтез гистоновых белков, которые принимают участие в формировании нуклеосомной структуры хроматина.
Существуют также сложные кластеры генов, которые могут кодировать длинные полипептиды с несколькими ферментативными активностями. Например, один из генов NeuraSpora grassa кодирует полипептид с молекулярной масой 150000 дальтон, который отвечает за 5 последовательных этапов в биосинтезе ароматических аминокислот. Полагают, что полифункциональные белки имеют несколько доменов – конформационно ограниченных полуавтономных образований в полипептидной цепи, выполняющих специфические функции. Открытие полуфункциональных белков дало основание полагать, что они являются одним из механизмов плейотропного действия одного гена на формирование нескольких признаков.
В кодирующей последовательности этих генов могут вклиниваться некодирующие, называемые интронами. Кроме того между генами могут находится участки спейсерной, и сателитной ДНК (рис.8.4).

Рис.8.4. Структурная организация нуклеотидных последовательностей (генов) в ДНК.
Спейсерная ДНК располагается между генами и не всегда транскрибируется. Иногда участок такой ДНК между генами (так называемый спейсер) содержит какую-то информацию, относящуюся к регуляции транскрипции, но он может представлять собой и просто короткие повторяющиеся последовательности избыточной ДНК, роль которой остается неясной.
Сателитная ДНК содержит большое количество групп повторяющихся нуклеотидов, которые не имеют смысла и не транскрибируются. Эта ДНК часто располагается в области гетерохроматина центромер митотических хромосом. Одиночные гены среди сателитной ДНК имеет регулирующие и усиливающие действие на структурные гены.
Большой теоритический и практический интерес для молекулярной биологии и медицинской генетики представляет микро- и минисателитные ДНК.
Микросателитная ДНК – короткие тандемные повторы из 2-6, (чаще из 2-4) нуклеотидов, которые получили название STR. Наиболее распространенными являются нуклеотидные ЦА- повторы. Количество повторов может существенно различатся у разных людей. Микросателиты находятся преимущественно в некоторых участках ДНК и насследуются по законам Менделя. Ребенок получают одну хромосому от матери, с определенным количеством повторов, другую от отца - с другим количеством повторов. Если рядом с геном ответственным за моногенное заболевание, или внутри гена расположен такой кластер микросателитов, то маркером патологического гена может быть определенное количество повторов по длине кластера. Эта особенность используется при непрямой диагностике генных болезней.
Минисателитная ДНК – тандемные повторы из 15-100 нуклеотидов. Они получили название VNTR – вариабельные по количеству тандемные повторы. Длина этих локусов также существено вариабельна у разных людей и может быть маркером (меткой) патологического гена.
Микро- и макросателитные ДНК используют:
1. Для диагностики генных болезней;
2. В судебно-медицинской экспертизе для идентификации личностей;
3. Для установления отцовства и в других ситуациях.
Наряду со структурными и регуляторными повторяющимися последовательностями, функции которых неизвестны, обнаружены мигрирующие нуклеотидные последовательности (транспозоны, мобильные гены), а также так называемые псевдогены у эукариот.
Псевдогены – нефункционирующие последовательности ДНК, которые сходные с функционирующими генами.
Вероятно, они произошли путем дупликации, а неактиаными копии стали в результате мутаций, нарушивших какие-либо стадии экспрессии.
По одной из версий псевдогены являются «эволюционным резервом»; по-другой – представляют собой «тупики эволюции», побочный эффект перестроек некогда функционирующих генов.
Транспозоны - структурно и генетически дискретные фрагменты ДНК, способные перемещаться от одной молекулы ДНК к другой. Впервые предсказаны Б.Мак-Клинток (рис. 8) в конце 40-х годов XX века на основе генетических экспериментов на кукурузе. Изучая природу окраски зерен кукурузы она сделала предположение, что существуют так званые мобильные („прыгающее”) гены, которые могут перемещаться по геному клетки. Пребывая по соседству с геном ответственным за пигментацию зерен кукурузы мобильные гены блокируют его работу. В дальнейшем транспозоны были выявлены у бактерий и было установлено, что они ответственны за устойчивость бактерий к различным токсическим соединениям.

Рис. 8.5. Барбара Мак- Клинток Впервые предсказала о существовании мобильных («прыгающих») генов, способных перемещаться по геному клеток.
Мобильные генетические элементы выполняют такие функции:
1. кодируют белки, ответственные за их перемещение и репликацию.
2. вызывают многие наследственные изменения в клетках, вследствии чего образуется новый генетический материал.
3. приводит к образованию раковых клеток.
4. встраиваясь в различные участки хромосом, они инактивируют или усиливают экспрессию клеточных генов,
5. является важным фактором биологической эволюции.
Современное состояние теории гена
Современные теории гена сформирована благодаря переходу генетики на молекулярный уровень анализа и отражает тонкую структурно-функциональную организацию единиц насследственности. Основные положения этой теории следующие:
1) ген(цистрон) – функциональная неделимая еденица наследственного материала (ДНК у организмов и РНК у некоторых вирусов), определяющая проявление наследственного признака или свойства организма.
2) Большинство генов существует в виде двух или большего числа альтернативных (взаимоисключающих) вариантов аллелей. Все аллели данного гена локализуются в одной и той же хромосоме в определенном ее участке, которую назвали локусом.
3) Внутри гена могут происходить изменения в виде мутаций и рекомбинаций; минимальные размеры мутона и рекона равны одной паре нуклеотидов.
4) Существуют структурные и регуляторные гены.
5) Структурные гены несут информацию о последовательности аминокислот в определенном полипептиде и нуклеотидов в рРНК, тРНК
6) Регуляторные гены контролируют и направляют роботу структурных генов.
7) Ген не принимает непосредственного участия в синтезе белка, он является матрицей для синтеза различных видов РНК, которые непосредственно принимают участие в синтезе белка.
8) Существует соответствие (колиннеарность) между расположением триплетов из нуклеотидов в структурных генах и порядком аминокислот в молекуле полипептида.
9) Большинство мутаций гена не проявляются в фенотипе, так как молекулы ДНК способны к репарации (востановлению своей нативной структуры)
10) Генотип являет собой систему, которая состоит из дискретных едениц – геннов.
11) Фенотическое проявление гена зависит от генотипической среды, в которой находится ген, влияние факторов внешней и внутренней среды.
В 1909 году датский ученый В. Йогансен предложил термин «ген». Ген – это участок ДНК хромосомы, который несет информацию об одном белке, молекуле транспортной или рибосомальной РНК. Функционально ген представляет собой целостную единицу. Но ген состоит из отдельных частей. Одни из которых активны – с них копируется РНК – экзоны . С других – не копируются. Они называются интронами . Количество интронов в генах разное, специфическое для каждого гена.
Структурные и регуляторные гены

Строение генов прокариот. Модель оперона, предложенная Ф. Жакобом и Ж. Л. Моно
Гены бывают структурные и регуляторные. Структурные – это гены, которые кодируют структуру рибонуклеиновых кислот и белков. Имеют большие размеры (сотни и тысячи нуклеотидов).
Регуляторные – это гены, которые служат местом присоединения ферментов или биологически активных соединений, которые влияют на активность структурных генов, принимают участие в процессах удвоения ДНК и транскрипции. Имеют незначительные размеры (до нескольких паров нуклеотидов).
Геномы и спейсеры

Строение генов эукариот
Количество генов у разных организмов разное. Совокупность генов в гаплоидном наборе, характерное для определенного вида, называется геномом .
Простейшими геномами являются геномы вирусов. Состоят лишь из структурных генов.
Геномы прокариот имеют и структурные, и регуляторные гены. Половина длины молекулы не несет информации, так как участки между генами – спейсеры – «разделительные знаки».
Геномы эукариот имеют наиболее сложную структуру. В них большее количество ДНК, поэтому много и структурных, и регуляторных генов. Например, у дрозофилы в геноме есть около 180 000 000 пар нуклеотидов, структурных генов – почти 10 тыс. В геномах организмов эукариот количество ДНК всегда значительно больше необходимого для кодирования (в 8-10 раз). Причины разные. Могут много раз повторяться последовательности нуклеотидов. В молекулах ДНК всегда много участков, которые не несут информации (спейсеры ).
"хромосома" - слова, которые знакомы каждому школьнику. Но представление об этом вопросе довольно обобщенное, так как для углубления в биохимические дебри требуются специальные знания и желание все это понимать. А оно, если и присутствует на уровне любопытства, то быстро пропадает под тяжестью изложения материала. Попробуем разобраться в хитросплетениях в научно-полярной форме.
Ген - это наименьшая структурная и функциональная частица информации о наследственности у живых организмов. По сути он представляет собой небольшой участок ДНК, в котором содержится знание об определенной последовательности аминокислот для построения белка либо функциональной РНК (с которой также будет синтезирован белок). Ген определяет те признаки, которые будут наследоваться и передаваться потомками дальше по генеалогической цепи. У некоторых одноклеточных организмов существует перенос генов, который не имеет отношения к воспроизведению себе подобных, он называется горизонтальным.
"На плечах" генов лежит огромная ответственность за то, как будет выглядеть и работать каждая клетка и организм в целом. Они управляют нашей жизнью от момента зачатия до самого последнего вздоха.

Первый научный шаг вперед в изучении наследственности был сделан австрийским монахом Грегором Менделем, который в 1866 году опубликовал свои наблюдения о результатах при скрещивании гороха. Наследственный материал, который он использовал, четко показывал закономерности передачи признаков, таких как цвет и форма горошин, а также цветки. Этот монах сформулировал законы, которые сформировали начало генетики как науки. Наследование генов происходит потому, что родители отдают своему чаду по половинке всех своих хромосом. Таким образом, признаки мамы и папы, смешиваясь, образуют новую комбинацию уже имеющихся признаков. К счастью, вариантов больше, чем живых существ на планете, и невозможно отыскать двух абсолютно идентичных существ.
Мендель показал, что наследст-венные задатки не смешиваются, а передаются от родителей потомкам в виде дискретных (обособлен-ных) единиц. Эти единицы, представленные у особей парами (аллелями), остаются дискретными и передаются по-следующим поколениям в мужских и женских га-метах, каждая из которых содержит по одной едини-це из каждой пары. В 1909 году датский ботаник Иогансен назвал эти единицы генами. В 1912 году генетик из Соединенных Штатов Америки Морган показал, что они находятся в хромосомах.
С тех пор прошло больше полутора веков, и исследования продвинулись дальше, чем Мендель мог себе представить. На данный момент ученые остановились на мнении, что информация, находящаяся в генах, определяет рост, развитие и функции живых организмов. А может быть, даже и их смерть.
Классификация
Структура гена содержит в себе не только информацию о белке, но и указания, когда и как ее считывать, а также пустые участки, необходимые для разделения информации о разных белках и остановки синтеза информационной молекулы.
Существует две формы генов:
- Структурные - они содержат в себе информацию о строении белков или цепей РНК. Последовательность нуклеотидов соответствует расположению аминокислот.
- Функциональные гены отвечают за правильную структуру всех остальных участков ДНК, за синхронность и последовательность ее считывания.
На сегодняшний день ученые могут ответить на вопрос: сколько генов в хромосоме? Ответ вас удивит: около трех миллиардов пар. И это только в одной из двадцати трех. Геном называется наименьшая структурная единица, но она способна изменить жизнь человека.
Мутации

Случайное или целенаправленное изменение последовательности нуклеотидов, входящих в цепь ДНК, называется мутацией. Она может практически не влиять на структуру белка, а может полностью извратить его свойства. А значит, будут локальные или глобальные последствия такого изменения.
Сами по себе мутации могут быть патогенными, то есть проявляться в виде заболеваний, либо летальными, не позволяющими организму развиваться до жизнеспособного состояния. Но большинство изменений проходит незаметно для человека. Делеции и дупликации постоянно совершаются внутри ДНК, но не влияют на ход жизни каждого отдельного индивидуума.
Делеция - это потеря участка хромосомы, который содержит определенную информацию. Иногда такие изменения оказываются полезными для организма. Они помогают ему защититься от внешней агрессии, например вируса иммунодефицита человека и бактерии чумы.
Дупликация - это удвоение участка хромосомы, а значит, и совокупность генов, которые он содержит, также удваивается. Из-за повторения информации она хуже подвержена селекции, а значит, может быстрее накапливать мутации и изменять организм.
Свойства гена
У каждого человека имеется огромная Гены - это функциональные единицы в ее структуре. Но даже такие малые участки имеют свои уникальные свойства, позволяющие сохранять стабильность органической жизни:
- Дискретность - способность генов не смешиваться.
- Стабильность - сохранение структуры и свойств.
- Лабильность - возможность изменяться под действием обстоятельств, подстраиваться под враждебные условия.
- Множественный аллелизм - существование внутри ДНК генов, которые, кодируя один и тот же белок, имеют разную структуру.
- Аллельность - наличие двух форм одного гена.
- Специфичность - один признак = один ген, передающийся по наследству.
- Плейотропия - множественность эффектов одного гена.
- Экспрессивность - степень выраженности признака, который кодируется данным геном.
- Пенетрантность - частота встречаемости гена в генотипе.
- Амплификация - появление значительного количества копий гена в ДНК.
Геном

Геном человека - это весь наследственный материал, который находится в единственной клетке человека. Именно в нем содержатся указания о построении тела, работе органов, физиологических изменениях. Второе определение данного термина отражает структуру понятия, а не функцию. Геном человека - это совокупность генетического материала, упакованного в гаплоидном наборе хромосом (23 пары) и относящегося к конкретному виду.
Основу генома составляет молекула хорошо известная как ДНК. Все геномы содержат по крайней мере два вида информации: кодированная информация о структуре молекул-посредников (так называемых РНК) и белка (эта информация содержится в генах), а также инструкции, которые определяют время и место проявления этой информации при развитии организма. Сами гены занимают небольшую часть генома, но при этом являются его основой. Информация, записанная в генах, — своего рода инструкция для изготовления белков, главных строительных кирпичиков нашего тела.
Однако для полной характеристики генома недостаточно заложенной в нем информации о структуре белков. Нужны еще данные об элементах которые принимают участие в работе генов, регулируют их проявление на разных этапах развития и в разных жизненных ситуациях.
Но даже и этого мало для полного определения генома. Ведь в нем присутствуют также элементы, способствующие его самовоспроизведению (репликации), компактной упаковке ДНК в ядре и еще какие-то непонятные пока еще участки, иногда называемые «эгоистичными» (то есть будто бы служащими только для самих себя). По всем этим причинам в настоящий момент, когда заходит речь о геноме, обычно имеют в виду всю совокупность последовательностей ДНК, представленных в хромосомах ядер клеток определенного вида организмов, включая, конечно, и гены.
Размер и структура генома

Логично предположить, что ген, геном, хромосома отличаются у разных представителей жизни на Земле. Они могут быть как бесконечно маленькими, так и огромными и вмещать в себе миллиарды пар генов. Структура гена также будет зависеть от того, чей геном вы исследуете.
По соотношению между размерами генома и числом входящих в него генов можно выделить два класса:
- Компактные геномы, имеющие не более десяти миллионов оснований. У них совокупность генов строго коррелирует с размером. Наиболее характерны для вирусов и прокариотов.
- Обширные геномы состоят более чем из 100 миллионов пар оснований, не имеющих взаимосвязи между их длиной и количеством генов. Чаще встречаются у эукариотов. Большинство нуклеотидных последовательностей в этом классе не кодируют белков или РНК.
Исследования показали, что в геноме человека находится около 28 тысяч генов. Они неравномерно распределены по хромосомам, но значение этого признака остается пока загадкой для ученых.
Хромосомы

Хромосомы - это способ упаковки генетического материала. Они находятся в ядре каждой эукариотической клетки и состоят из одной очень длинной молекулы ДНК. Их легко можно увидеть в световой микроскоп в процессе деления. Кариотипом называется полный набор хромосом, который является специфичным для каждого отдельного вида. Обязательными элементами для них являются центромера, теломеры и точки репликации.
Изменения хромосом в процессе деления клетки
Хромосома - это последовательные звенья цепи передачи информации, где каждое следующее включает предыдущее. Но и они претерпевают определенные изменения в процессе жизни клетки. Так, например, в интерфазе (период между делениями) хромосомы в ядре расположены рыхло, занимают много места.
Когда клетка готовится к митозу (т. е. к процессу разделения надвое), хроматин уплотняется и скручивается в хромосомы, и теперь его становится видно в световой микроскоп. В метафазе хромосомы напоминают палочки, близко расположенные друг к другу и соединенные первичной перетяжкой, или центромерой. Именно она отвечает за формирование веретена деления, когда группы хромосом выстраиваются в линию. В зависимости от размещения центромеры существует такая классификация хромосом:
- Акроцентрические - в этом случае центромера расположена полярно по отношению к центру хромосомы.
- Субметацентрические, когда плечи (то есть участки, находящиеся до и после центромеры) неравной длины.
- Метацентрические, если центромера разделяет хромосому ровно посередине.
Данная классификация хромосом была предложена в 1912 году и используется биологами вплоть до сегодняшнего дня.
Аномалии хромосом

Как и с другими морфологическими элементами живого организма, с хромосомами тоже могут происходить структурные изменения, которые влияют на их функции:
- Анеуплоидия. Это изменение общего числа хромосом в кариотипе за счет добавления или удаления одной из них. Последствия такой мутации могут быть летальными для еще не родившегося плода, а также приводить к врожденным дефектам.
- Полиплоидия. Проявляется в виде увеличения количества хромосом, кратного половине их числа. Чаще всего встречается у растений, например водорослей, и грибов.
- Хромосомные аберрации, или перестройки, - это изменения в строении хромосом под воздействием факторов внешней среды.
Генетика
Генетика - это наука, изучающая закономерности наследственности и изменчивости, а также обеспечивающие их биологические механизмы. В отличие от многих других биологических наук она с момента своего возникновения стремилась быть точной наукой. Вся история генетики — это история создания и использования все более и более точных методов и подходов. Идеи и методы генетики играют важную роль в медицине, сельском хозяйстве, генетической инженерии, микробиологической промышленности.
Наследственность - способность организма обеспечивать в ряду морфологических, биохимических и физиологических признаков и особенностей. В процессе наследования воспроизводятся основные видоспецифические, групповые (этнические, популяционные) и семейные черты строения и функционирования организмов, их онтогенеза (индивидуального развития). Наследуются не только определенные структурно-функциональные характеристики организма (черты лица, некоторые особенности обменных процессов, темперамента и др.), но и физико-химические особенности строения и функционирования основных биополимеров клетки. Изменчивость — разнообразие признаков среди представителей определенного вида, а также свойство потомков приобретать отличия от родительских форм. Изменчивость вместе с наследственностью представляют собой два неразделимых свойства живых организмов.
Синдром Дауна

Синдром Дауна - генетическое заболевание, при котором кариотип состоит из 47 хромосом у человека вместо обычных 46. Это одна из форм анеуплоидии, о которой говорилось выше. В двадцать первой паре хромосом появляется добавочная, которая привносит лишнюю генетическую информацию в геном человека.
Название свое синдром получил в честь врача, Дона Дауна, который открыл и описал его в литературе как форму психического расстройства в 1866 году. Но генетическая подоплека была обнаружена почти на сто лет позже.
Эпидемиология

На данный момент кариотип в 47 хромосом у человека встречается один раз на тысячу новорожденных (ранее статистика была иной). Это стало возможным благодаря ранней диагностике данной патологии. Заболевание не зависит от расы, этнической принадлежности матери или ее социального положения. Оказывает влияние возраст. Шансы родить ребенка с синдромом Дауна возрастают после тридцати пяти лет, а после сорока соотношение здоровых детей к больным равняется уже 20 к 1. Возраст отца старше сорока лет также увеличивает шансы на рождение ребенка с анеуплоидией.
Формы синдрома Дауна
Наиболее частый вариант - появление дополнительной хромосомы в двадцать первой паре по ненаследственному пути. Он обусловлен тем, что во время мейоза эта пара не расходится по веретену деления. У пяти процентов заболевших наблюдается мозаицизм (дополнительная хромосома содержится не во всех клетках организма). Вместе они составляют девяносто пять процентов от общего количества человек с этой врожденной патологией. В остальных пяти процентах случаев синдром вызван наследственной трисомией двадцать первой хромосомы. Однако рождение двух детей с этим заболеванием в одной семье незначительно.
Клиника
Человека с синдромом Дауна можно узнать по характерным внешним признакам, вот некоторые из них:
Уплощенное лицо;
- укороченный череп (поперечный размер больше продольного);
- кожная складка на шее;
- складка кожи, которая прикрывает внутренний угол глаза;
- чрезмерная подвижность суставов;
- сниженный тонус мышц;
- уплощение затылка;
- короткие конечности и пальцы;
- развитие катаракты у детей старше восьми лет;
- аномалии развития зубов и твердого неба;
- врожденные пороки сердца;
- возможно наличие эпилептического синдрома;
- лейкозы.
Но однозначно поставить диагноз, основываясь только на внешних проявлениях, конечно, нельзя. Необходимо провести кариотипирование.
Заключение
Ген, геном, хромосома - кажется, что это просто слова, значение которых мы понимаем обобщенно и весьма отдаленно. Но на самом деле они сильно влияют на нашу жизнь и, изменяясь, заставляют меняться и нас. Человек умеет подстраиваться под обстоятельства, какими бы они ни оказались, и даже для людей с генетическими аномалиями всегда найдется время и место, где они будут незаменимы.